

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
In adults, the incisive canal is a narrow bony canal connecting the nasal cavity and incisive fossa, allowing passage of the nasopalatine nerve, a branch of the greater palatine artery and, in some individuals, a remnant of the nasopalatine duct [1]. The incisive canal is generally regarded as passing through the border between the primary and secondary palates, i.e., along the incisive-maxillary or transverse suture of the bony palate [2,3,4]. Gray's Anatomy [1] also described the course along the bony suture. The bony canal in adults consistently opens to an incisive fossa at a cross point of the transverse and longitudinal sutures of the bony palate. Moreover, examination of sagittal sections showed the presence of the duct, nerves and vessels through a canal along the suture [3567]. Therefore, the canal is usually considered to pass through a border between the primary and secondary palates, while the duct is commonly known as a derivative from a primary oronasal communication. However, Vacher et al. [3] demonstrated that, in sagittal sections of a single fetus at 14 weeks, the canal passes through the incisive bone (IN) or premaxilla. Moreover, Radlanski et al. [8] postulated a new concept that the canal develops within the embryonic IN or primary palate according to a relatively small number of specimens after palatal closure (six fetuses at approximately 10–27 weeks). Could such a limited observation change one of essential knowledges of embryology? Otherwise, are there individual and/or age-related variations present in topographical relation between the incisive canal and suture?
Using frontal sections of human midterm fetuses, our group recently demonstrated that the bony canal penetrates the IN, not along the IN-maxillary suture, in fetuses “with” a persistent nasopalatine duct [9]. It is unclear, however, whether the intra-IN course of the duct corresponds to a bony variation or anomaly. Is the intra-IN canal likely when the fetuses had no epithelial duct? In contrast to our previous study using frontal sections [9], comprehensive studies of the bony palate (e.g., Wood and Kraus [10]) were almost always based on observations of sagittal sections. However, since a single sectional plane chosen is likely to make an identification of thin bony structures difficult, we need to compare results from multiple sectional planes. Consequently, in spite of the long history of research, we should emphasize that anatomy of the incisive canal and duct is still debatable. This study therefore assessed the topographical anatomy of the incisive canal and the IN-maxillary suture using a large number of midterm and latestage fetuses with or without a persistent nasopalatine duct.
Materials and Methods
This study was performed in accordance with the principles of the Declaration of Helsinki 1995 (as revised in 2013). Serial or semiserial sections were obtained from 69 fetuses, including 31 specimens at gestational age (GA) 9–15 weeks and crown rump length (CRL) 36—110 mm and 38 of GA 26–34 weeks and CRL 210–290 mm. The sectional planes were sagittal in 32 specimens (nine midterm and 23 late-stage) and frontal in 37 specimens (22 midterm and 15 late-stage). In the world, to our knowledge, there is no collection of histological sections from abundant fetuses at 20–25 weeks or CRL 140–200 mm.
The 31 specimens of midterm fetuses were part of the large collection kept at the Embryology Institute of the Universidad Complutense, Madrid, and originated from miscarriages and ectopic pregnancies at the Department of Obstetrics of the University. All specimens were sectioned serially (frontal or sagittal) (Table 1). Most sections were stained with hematoxylin and eosin, with others stained with Masson trichrome, azan, orange G, or silver stain. The study protocol was approved by the ethics committee of the Universidad Complutense (B08/374). All midterm specimens were of Spanish white origin.
The 38 specimens of late-stage fetuses were parts of the collection of the Department of Anatomy, Akita University, Akita, Japan. These specimens had been donated by their families to the Department from 1975—1985 and preserved in 10% (w/w) neutral formalin solution for more than 30 years. The available data were limited to the date of donation and GA, but there was no information on family name, the name of the obstetrician or hospital and the reason for abortion. All late-stage specimens were of Far-East yellow origin, most likely of Japanese. The use of these specimens for research was approved by the ethics committee of Akita University (No. 1428). After removal of the palate in combination with its surrounding tissues including lower parts of the nose and the pterygoid of the sphenoid, the specimens were decalcified by incubating them at room temperature in Plank-Rychlo solution (AlCl2/6H2O, 7.0 w/v%; HCl, 3.6; HCOOH, 4.6) for 1–2 weeks. After routine procedure for paraffin embedded histology, frontal or sagittal semiserial sections (numbers of specimen) (Table 1), 5-µm-thick at 50-µm intervals, were prepared from each block and stained with hematoxylin and eosin. All observations and photographs were made with a Nikon Eclipse 80 camera (Nikon, Tokyo, Japan).
Results
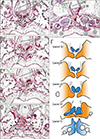
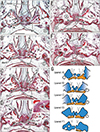
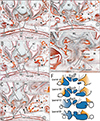
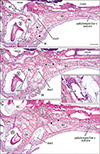
Figs. 1, 2, 3, 4 display bony palates from midterm fetuses, while Figs. 5, 6, 7 show bony palates from late-stage fetuses, with frontal sections shown in Figs. 1, 2, 3 and 5 and sagittal sections in Figs. 4, 6, and 7. Frontal sections, of which posterior site is always shown in the top panel, were better at showing the superomedial processes of the IN. Likewise, frontal sections were able to show a fact that an essential left/right difference was absent in the present fetuses for them. Although Figs. 3E and 5F might make an impression of the left-right difference, a same bony structure was seen in different anteroposterior levels (e.g., Fig. 5D vs. Fig. 5F). Below, we will describe three types of the incisive canal or possible individual variations: a correspondence between a figure and the type is summarized in Table 2.
Superomedial processes of the IN
In frontal sections, the bony processes were consistently paired and attached to each other across a narrow midline suture below the vomer. Thus, midline sutures of bilateral INs were observed not only in the lower large mass near the oral cavity but also between the upper processes: both were likely to be included in a single frontal section (Figs. 2C, 3E, 5I). These processes were adjacent to anterior (alar) nasal cartilages at anterior sites (Figs. 1B, 2C, 3D) as well as to the vomers at posterior sites (Figs. 3A, B, 5C, F, I). At anterior sites, the INs had generated plate-like processes similar to the paired processes of the vomer sandwiching the nasal septum (Figs. 1E, 5I). Likewise, the maxilla was also likely to issue a plate-like anterior part lining the inferior or medial aspect of incisive canal (IC) (Figs. 2E, F, 5B, F, I). These process and plate provided a complicated anatomy of a border or suture between the IN and maxilla.
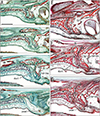
In sagittal sections, the aforementioned superomedial process of the IN was sometimes identified as a single plate (Fig. 4B) but often identified as fragments (Figs. 4F, 7A), making identification of the IN-maxillary suture more difficult than in frontal sections. Moreover, the apparent branching or joining of the suture and/or canal in sagittal sections (Figs. 4F, 7B) also made understanding difficult, inasmuch as the suture branches were consistently located on the posterior side of the incisive canal.
Incisive-maxillary suture and the incisive canal
At sites distant from the incisive canal, the IN-maxillary sutures were narrow and tight and contained no or few vessels (stars in Figs. 2B, 3B, 4A, E, 5A, D, G, 6A, 7A). Sutures along the lateral aspect of the IN were difficult to identify because they consisted only of loose mesenchymal tissue without trabeculae (stars in Figs. 1D, 2A right, 3D right). In contrast, the incisive canal was always thick and contained vessel-rich loose tissue continuous with a submucosal layer of the nose, a histological similarity easily identifying the incisive canal, irrespective of whether the section included an elongation of nasal epithelium (see below). Frontal sections, with a wide view of the nasal cavity, clearly demonstrated the continuation of tissue from the nose to the canal (Figs. 1C, 2A, 3B, 5B, F, H). In contrast, sagittal sections along and near the midline showed that the vomer and nasal septum cartilage occupied a large area above the upper opening of the incisive canal (Figs. 4D, 7A). Frontal sections of the course almost along the left-right axis also clearly showed the inferomedial elongation of the nasal epithelium entering the incisive canal (Figs. 1A, B, 2A, 3A, 5B, F, H). This epithelial elongation sometimes reached an epithelial pearl in the incisive fossa (Figs. 2E, 4G, 5C), a morphology defined as being a real nasopalatine duct [911]. This morphology was observed in 22 fetuses of CRL >70 mm or GA 11 weeks.
Intra-incisive canal and the canal along the suture
The development of the incisive canal, corresponding to the IN-maxillary suture, resulted in partial enlargement of the thin and tight sutures, which contained vessel-rich loose tissue in 23 of the 69 fetuses (33.3%) (Figs. 1B, 4C, F, 5B, 7B). Along the canal, a thin layer of the maxilla sometimes covered the medial aspect of the IN (Fig. 5B, C). However, other than the maxilla, the incisive canal in 46 fetuses (66.7%) was sandwiched partly or entirely by the upper process and the lower mass of the IN. In 35 of these fetuses, the canal penetrated the IN at the nasal half of the course (Figs. 2B, C, 5E, F, 7A, B), whereas, in the other 11, the canal penetrated the IN along the entire course (Figs. 3C, 5I). The course of the canal was not connected with the presence of a persistent nasopalatine duct. Topographical variations between the canal and suture at midterm are summarized in Figs. 1F, 2G, and 3F. In addition, candidate nasopalatine nerves were observed in (1) the incisive canal and incisive fossa (Figs. 3B, E, 5B, E, H), (2) the midline suture of the bilateral INs (Figs. 1D, 2D), and/or (3) the center of the lower mass of the IN on the superoposterior side of the second tooth (Figs. 2D, 4C, E, 6C, D, 7A, B). These nerves differed from the branches of the major palatine nerve coming from the posterior site (Fig. 6A, B).
Bias suggested by the sectional plane and other factors
Although sagittal sections were advantageous for understanding the anteroposterior arrangement of the IN, maxilla, palatal bone and upper teeth, the vomer and nasal septum cartilage of late-stage specimens tended to be injured during histological preparation of sagittal sections (Fig. 7). Because of difficulties identifying irregularly shaped upper processes of the IN and because of the course of the incisive canal along the left-right axis near the nose, the intra-IN canal tended to be more frequently observed in frontal than in sagittal sections (Table 1). Sagittal sections sometimes provided a complicated morphology, in that the IN-maxillary suture appeared to be a “branch” of the incisive canal (Figs. 4F, 7B, C). Our observations of late-stage specimens were not based on serial sections but on semiserial sagittal sections at 50 micron intervals. We were concerned that this method may be associated with a failure to identify the small upper processes of the IN. However, possibly due to easy identification of thick loose tissue continuous with the submucosa of the nose, the intra-IN canal tended to be found more frequently in semiserial sections of late-stage fetuses than in serial sections of midterm fetuses (Table 1). There was no evidence of rearrangement or reconstruction of the upper parts of the IN during intrauterine growth after 15 weeks.
Consequently, in both of midterm and late-stage fetuses, we found not only the intra-IN canal but also the “classical” canal along the IN-maxillary suture (Figs. 1, 4, 6). There were variations between both of the typical morphologies (“mixed” in Table 2). Nevertheless, we should pay much attention to avoid bias from sectional planes. Sagittal sections were better for the demonstration of the classical canal because, conversely, frontal sections emphasized upper details of the IC as well as a plate-like anterior part of the maxilla.
Discussion
A study history of the incisive canal is very long and, still now, the structure is generally considered as a product according to one of basic rules of embryology, i.e., a gap of the border between the primary and secondary palates (see the Introduction). However, the present observation demonstrated that the incisive canal with or without nasopalatine duct exhibited a spectrum of variations in topographical relation to the IN-maxillary suture. In contrast to the recent new concept [8], some fetuses carried the intra-IN canal opening to the incisive fossa, while the others had the incisive canal along the IN-maxillary suture.
We identified an intra-IN canal, with or without a nasopalatine duct, as a normal morphology. The upper half of each incisive canal was characterized by both the inferomedial elongation of the nasal epithelium and a continuation of the thick nasal submucosal tissue. The canal seemed to be formed during nasal epithelial development at midterm. Conversely, the development of the intra-IN canal did not require a persistent nasopalatine duct reaching the incisive fossa or connecting with the epithelial pearl. Thus, this study did not emphasize our previous definition of the nasopalatine duct as a duct connecting the nasal epithelium and the paramedian epithelial pearl [9]. This duct apparently did not originate from the primitive oronasal communication prior to the midline palatal fusion. Although we previously reported the presence of this duct in specimens of GA 14 weeks [9], in the present study we identified this duct in specimens of GA 11 weeks. This lower limitation of age or stage was difficult to explain if the duct originated from the primitive oronasal communication during early phases of palatal fusion.
Some cells packed in the midline epithelial seam after fusion may later form a duct that connects to the differentiated nasal epithelium. Actually, the midline seam connects to the epithelial pearls in the incisive fossa [111213]. However, the nasopalatine duct in the incisive canal often penetrated the IN, not the IN-maxillary suture. The primitive oronasal communication passes through the suture, as the latter was apparently the most anterior site in the loose midline. Therefore, conversely, most or all nasopalatine ducts likely originate from the “secondary” elongation of the nasal epithelium after midterm. The irregularly shaped upper parts of the IN with several processes seemed also to be a product of invasion of the nasal epithelium and submucosal tissue.
Previous studies showed “sagittal sections” that included the incisive canal along the IN-maxillary suture [2567, 14], but these studies did not mention the possibility of an intra-IN canal, which for a long time had not been a focus of study. Thus, at the beginning of this study, we considered that a midterm fetus with an intra-IN canal would likely die before reaching late-stage or full term. Indeed, this study proofed that fetuses with the intra-IN canal could be alive until late-stage or even full term. However, sagittal sections themselves had disadvantage for the demonstration. First, it might be difficult to define the margins of the vomer and anterior nasal cartilages adjacent to the IN during fetal development [415161718]. Second, the upper processes of the IN were difficult to identify on sagittal sections. In most samples, the intra-IN canal was observed in the nasal half of the course, lying between the upper process and the lower large mass of the bone. Third, an inferomedial elongation of the nasal epithelium, a simple marker for identifying the incisive canal, was found to run along the left-right axis rather than the supero-inferior axis. Therefore, the nasal epithelium entering the canal or suture was often not seen on sagittal sections. In contrast, frontal sections clearly demonstrated that the vomer was sandwiched between the nasal septum cartilage and the upper process of the IN.
Because the present specimens included large fetuses near term, newborns and children are also likely to carry the intra-IN canal. Advances in computed tomography have revealed various abnormalities of the incisive canal and nasopalatine duct in adults [192021222324]. These abnormalities were apparently caused by a remnant of a nasopalatine duct or its fragment, with or without upper migration, as these variants were connected to the nasal epithelium. However, a large incisive fossa along the midline on the oral surface of the palate might make a macroscopic finding of the intra-IN canal difficult. Detailed examinations of adult morphology may confirm the high incidence of intra-IN ducts. However, the topographical relationship between the IN and maxilla (i.e., the anatomy of the suture) may change during postnatal growth. In this context, we should remind a plate-like anterior part of the maxilla that lined the inferior or medial aspect of IN and that disturbed our clear identification of the IN-maxillary suture (Figs. 2E, F, 5B, F, I). We did not deny a possibility that, depending on eruption and growth of the face, these thin bony plates could be absorbed because they were located closely to anterior teeth and nose.
Study limitations
The present specimens were of two different origins: Spanish for the midterm and Far-East origin for the late-stage. Although this racial difference was likely to influence incidence of variations (Table 1), we were able to demonstrate a real presence of variations in one of the basic and famous structures in the human head. Due to the large size, we used semiserial sections (50-µm interval) for observations of the late-stage specimens. We believe that, because of more than 100-µm thicknesses of the canal and duct, even semiserial sections provided a precise information as to whether these structures were present or absent.



 XML Download
XML Download