

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
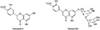
Hesperidin (4′-methoxy-7-O-rutinosyl-3′,5-dihydroxyflavanone; hesperetin 7-O-rutinoside) is a naturally occurring flavanone glycoside found in citrus [12]. Hesperidin is hydrolyzed by the intestinal microflora when ingested orally and then absorbed in the large intestine, and hesperetin (4′-methoxy-5,7,3′-trihydroxyflavanone), aglycone form of hesperidin, is mainly absorbed in the small intestine [3]. Hesperidin exerts immunoregulatory properties via the modification of lymphocyte composition in the intestinal mucosa [4] and in gut-associated lymphoid tissues [5]. The chemical structures of both hesperidin and hesperetin (Fig. 1) have been well-described in previous studies [36].
Due to its antioxidant and anti-inflammatory properties, hesperidin possesses a variety of biological effects in models of cardiovascular disease [78] and diabetes [29] as well for the prevention of cancer [8]. Additionally, the antioxidant and anti-inflammatory effects of hesperidin ameliorate symptoms in animal models of the neurodegenerative diseases of Alzheimer's disease [10], Parkinson's disease [1112], Huntington's disease [1314], depression [1516], neuroimmunological multiple sclerosis (MS) [1718], brain ischemia-reperfusion injury [19], and traumatic injury in central nervous system (CNS) tissues [20]. Although the etiologies of the abovementioned neuroinflammatory diseases are varied and distinct, oxidative stress and inflammatory responses are common and consistent findings in CNS tissues affected by these disorders. However, these changes are occasionally reversed by antioxidant flavonoids.
The beneficial effects of citrus flavonoids, including hesperidin, have been reviewed in the context of their neuropharmacological properties [3] and neuroprotective capabilities [212223] with a special focus on anti-depressive actions and protection against learning and memory deficits. Moreover, updated data on the effects of hesperidin in animal models of neurological disorders remain to be further analyzed. The present review will provide updates on recent publications assessing the effects of hesperidin in models of oxidative stress-related neuroinflammation including lipopolysaccharide (LPS)-induced endotoxemia, brain ischemia-reperfusion injury, and chemical-induced hippocampal dysfunction. Because the etiologies of autoimmune CNS diseases are distinct from those of other neurodegenerative diseases, this review will discuss the potential role that hesperidin plays in autoimmune diseases.
For this review, the following terms were searched for in the PubMed database: “hesperidin and antioxidant and inflammation” (79 articles); “hesperidin and central nervous system” (66 articles); “hesperidin and autoimmune” (9 articles); and “hesperidin and demyelination” (5 articles). For the data extraction, all authors of the present review determined the eligibility of the articles.
Molecular Mechanisms of Hesperidin (Antioxidation and Anti-inflammation)
The antioxidant activities of hesperidin have been previously reported [17]. Briefly, the beneficial effects of hesperidin are largely dependent on its radical scavenging activities and its augmentation of cellular antioxidant mechanisms [7]. In terms of scavenging radicals, hesperidin restores deficits in the activity of antioxidant enzymes, including glutathione peroxidase, glutathione reductase, catalase, and superoxide dismutase, that are downregulated in the brain in experimental models of stroke [24], irradiation [25], and LPS-induced endotoxicity [26].
Similar to its antioxidant effects, hesperidin also activates cellular protection machinery, including nuclear factor-erythroid 2-related factor-2 and heme oxygenase-1, which are centrally involved in cell survival against oxidative stress [2728]. Regarding the signal cascades associated with cell protection, hesperidin stimulates peroxisome proliferator-activated receptor-gamma, which is centrally involved in the mediation of anti-inflammatory and antioxidant effects in models of stress-induced gastric ulcers [29], ischemic heart disease [30], and cyclophosphamide-induced hepatotoxicity [31].
The anti-inflammatory activities of hesperidin are indispensable to its antioxidant activities [7]. For example, hesperidin reduces the production of pro-inflammatory mediators, such as tumor necrosis factor α (TNF-α), in a model of aluminum chloride-induced neuroinflammation in the hippocampus [32] and in N-methyl-D-aspartate-induced excitotoxicity in the retina [33]. Furthermore, intraperitoneal injections of hesperidin to normal animals decrease the level of phosphorylated extracellular signal-regulated kinase-1 and -2, which are important signals in cell activation in the cerebral cortex, cerebellum, and hippocampus [34]. Thus, it can be postulated that, similar to other antioxidant flavonoids, hesperidin activates radical scavenging activity and subsequently attenuates the inflammatory response to be ultimately protective against cell death.
Bioactivity of Hesperidin in the Nervous System
Effects of hesperidin on neuroinflammation
In animal models, LPS triggers neuroinflammation in conjunction with the development of cognitive deficits and septic shock [26]. Additionally, LPS-induced neuroinflammation in mice is associated with upregulated levels of the mediators of oxidative stress and inflammation, including TNF-α, whereas hippocampal acetylcholinergic activity decreases due to the activation of acetylcholinesterase [26]. Hesperidin reverses the phenomena induced by LPS treatment by increasing the generation of radical scavenging enzymes, including superoxide dismutase and catalase. In LPS-treated animals, hesperidin also reduces the level of pro-inflammatory cytokines, including interleukin (IL)-1β, IL-6, and TNF-α, in the brain by promoting activation of the miRNA-132 pathway [35]. Even though the cellular sources of pro-inflammatory cytokines in LPS-induced neuroinflammation models have yet to be validated, it has been postulated that neuroglial cells, including microglia and astrocytes, are potential sources of these mediators.
In terms of the effects of hesperidin on neuroglial cells, a conditioned medium from hesperidin-treated astrocytes increases neuronal progenitor cells [36], which suggests that hesperidin-exposed astrocytes indirectly influence neuronal cell differentiation. Regarding the permeability of hesperidin through the blood-brain barrier, hesperidin may be absorbed in the large intestine due to its turnover to hesperetin via microflora [3] and then enter the blood circulation to, ultimately, penetrate the brain through the blood-brain barrier [37]. The production of nitric oxide and pro-inflammatory mediators, including TNF-α and IL-1β, are significantly suppressed by hesperidin in the RAW246.7 macrophage cell line [38]. Thus, there is a consensus that hesperidin suppresses the biological activity of astrocytes, microglia, or both, in multiple ways.
Taken together, these data suggest that hesperidin interferes with the initiation of inflammation in non-neuronal systems as well as the brain, possibly, via reductions in proinflammatory cytokines, enhancing the activation of radical scavenging enzymes, and restoring neuronal activity.
Effects of hesperidin on demyelinating diseases in the CNS
Effects of hesperidin in autoimmune demyelinating diseases
The types of neuroinflammation associated with autoimmune demyelinating diseases, such as MS and its animal models, as well as experimental autoimmune encephalomyelitis (EAE) are distinct from that of neurodegenerative diseases, such as Alzheimer's disease [39]. EAE is characterized by the generation of autoimmune T cells in the peripheral lymphoid tissues and the infiltration of these autoimmune T cells into the CNS parenchyma, which result in demyelination and reactive gliosis [40]. Thus, the suppression of T-cell proliferation during the induction stage of EAE is a primary target for the prevention of this disease. Additionally, the interruption of inflammatory cell migration into CNS tissues is another key step in preventing damage to CNS cells, including glial cells and neurons. Finally, the suppression of oxidative stress and inflammatory mediators might be an alternative method that can inhibit EAE progression [41]. Blocking at least one of these three processes would be applicable for the amelioration of EAE. Previous studies have shown that the antioxidant apigenin, which is a natural flavonoid, inhibits the progression of EAE via one of the processes described above [42].
In models of autoimmune CNS diseases, hesperidin improves EAE-induced paralysis when administered orally (50, 100, and 200 mg/kg [18]) or via subcutaneous injection (50 mg/kg/day for 7 consecutive days [17]). The mechanisms underlying the effects of hesperidin on EAE are largely dependent on its strong antioxidant and anti-inflammatory properties [1718]. Furthermore, hesperidin is involved in the suppression of autoimmune T cell proliferation, the activation of regulatory T cells, reductions in microglial cells in EAE lesions, and decreased demyelination [18], which could possibly lead to the M2 phenotype macrophage bias in EAE [41]. Thus, the beneficial effects of hesperidin on EAE appear to be largely dependent on suppressing the inflammatory response and oxidative stress in the peripheral immune system and EAE target tissues, which would lead to reductions in demyelination.
However, a study investigating the effects of hesperetin, which is a flavanone and an aglycone of hesperidin, reported that hesperetin treatment (approximately 10 mg/day/each in drinking water) delayed the recovery of paralysis in mice with proteolipid protein-induced EAE [43], which suggests that hesperetin has a detrimental effect in EAE. It is possible that the differential effects of hesperidin-related compounds on EAE depend on the treatment dose and different immunization antigens (myelin oligodendrocyte glycoprotein vs. proteolipid protein) because it is widely accepted that flavones, including luteolin and apigenin, inhibit antigen-specific proliferation and interferon-gamma production by murine and human autoimmune T cells in vitro [51].
Regarding the role of radicals in models of autoimmune disease, nitric oxide occasionally suppresses the proliferation of T cells and also induces cell apoptosis [52]. Therefore, reductions in nitric oxide levels may facilitate the proliferation of autoimmune T cells and interfere with the apoptotic elimination of inflammatory cells in target organs. This finding is further supported by studies that assessed treatment of the nitric oxide carrier in EAE [5354]. Thus, the hesperidin-induced reduction of radicals might be case-sensitive and/or inflammation stage–dependent in models of autoimmune CNS diseases.
Effects of hesperetin in a model of chemical-induced demyelination
Demyelination can be visualized via decreases in myelin, which can be caused by either the degradation of myelin-forming oligodendrocytes or direct myelin loss without the loss of oligodendrocytes [55]. A model of lysolecithin-induced focal demyelination [56] showed that this process is distinct from that of autoimmune demyelination in EAE, which is associated with the infiltration of inflammatory cells into the CNS parenchyma [41]. Lysolecithin-induced demyelination is characterized by myelin loss and glial activation without the infiltration of hematogenous cells and can be ameliorated by hesperetin treatment [57]. The beneficial effects of hesperetin (in the present review article, the term “hesperetin” will be used rather than “hersperitin” according to the generic name of the structural formula of the compound) have been attributed to its direct cytoprotective and antioxidant effects in CNS tissues, which is similar to the ability of the flavonoid quercetin regarding the repair of myelin [58]. This finding suggests that hesperetin can permeate through the bloodbrain barrier to influence neurons and neuroglial cells directly in damaged regions.
Effects of hesperidin in hippocampal dysfunction
The hippocampus is centrally involved in cognitive and memory functions as well as the progression of MS and the animal model of EAE. Currently, there is a consensus that MS patients suffer from cognitive dysfunction [5960] and that EAE mice exhibit hippocampal atrophy in conjunction with increased radical activity and the activation of microglial cells [6162]. In models of EAE, hesperidin exerts neuroprotective and other beneficial effects that, possibly, occur through the suppression of microglial cells. It is known that hesperidin protects neurons against death and promotes the survival of neuronal progenitor cells through activation of the phosphatidylinositol 3-kinase and mitogen-activated protein kinase pathways [63]. Even though the biochemical activity of neurons in models of EAE has yet to be precisely evaluated following hesperidin treatment, it is possible that hesperidin may protect neurons against glutamate excitotoxicity in EAE-affected brains [64], as it does in models of neurodegeneration [322].
The neuroprotective effects of hesperidin against hippocampal excitotoxicity were assessed in a model of aluminum chloride-induced excitotoxicity that leads to cell death with the concurrent hyperactivation of microglial cells [65]. The cytoprotective effects of hesperidin are largely dependent on its antioxidant and anti-inflammatory activities [3247], which indicates that cognitive impairments are ameliorated by hesperidin treatment. Hesperidin treatment is also associated with decreased levels of nitrate/nitrite and increased levels of brain-derived neurotrophic factor in the mouse hippocampus [45] as well as the inhibition of glutamate release following kainic acid-induced excitotoxicity in the rat hippocampus [49]. Furthermore, hesperidin improves memory consolidation in mice with streptozotocin-induced cognitive impairments by modulating acetylcholinesterase activity [48] and protects neurons against cell death [63]. Taken together, these findings indicate that hesperidin exerts protective effects in the hippocampus.
As a model of stroke, brain ischemia-reperfusion injury is characterized by cell loss and reactive gliosis in the hippocampus of adult [24] and neonatal [50] rats. However, hesperidin significantly reverses reductions in the activities of antioxidant enzymes, such as glutathione peroxidase, glutathione reductase, catalase, and superoxide dismutase, and ameliorates glutathione content following middle cerebral artery occlusion. Furthermore, hesperidin reduces the production of various inflammatory mediators, including TNF-α and IL-1β, and the expression of inducible nitric oxide synthase. Thus, it can be postulated that hesperidin-induced neuroprotection is partly dependent on the suppression of inflammatory responses in the hippocampus after middle cerebral artery occlusion [1924] and that hesperidin contributes to neuronal survival [63].
Effects of hesperidin on depressive-like behaviors
Hesperidin may also have beneficial effects on depressivelike behaviors because a majority of studies have found that treatment with hesperidin ameliorates depression, irrespective of the causative agent [3]. For example, hesperidin reduces immobility time in the forced swimming test [4445] and, possibly, promotes neuronal activity in the hippocampus. Additionally, a recent study reported that the mechanisms underlying the antidepressant-like effects of hesperidin include the modulation of serotonergic 5-HT1A receptors [46] and kappaopioid receptors in the hippocampus [44]. The neuroprotective activity and modulation of serotonergic neurons following hesperidin treatment also alter a variety of depressive-like behaviors in LPS-injected mice with endotoxemia [35], mice with traumatic brain injury [20], streptozotocin-induced diabetic rats [16], a 6-hydroxydopamine model of Parkinson's disease [12], and mice following an olfactory bulbectomy [15]. Thus, it can be postulated that the antidepressant-like effects of dietary citrus containing hesperidin are non-specifically obtained via a broad spectrum of neuroprotection, as described above [32223].
Conclusion
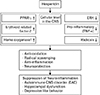
In general, the molecular mechanisms that underlie the beneficial effects of the flavone glycoside hesperidin on CNS disease remain unclear. However, citrus is known to exert positive effects because it contains a variety of non-toxic antioxidants. Decades of research on these antioxidant activities have provided experimental evidence showing that hesperidin originating from citrus plays an important neuroprotective role through its antioxidant and anti-inflammatory activities (Table 1). Moreover, the molecular mechanisms of hesperidin, underlying it effects in nervous tissues and immune cells, are largely dependent on the suppression of cell activation signals and radical scavenging effects (Fig. 2). In terms of nutraceutical effects, citrus without toxic effects can provide beneficial effects by preventing and curing CNS disorders, independently of inflammation type, in different lesions of CNS tissues. Therefore, the antioxidant and anti-inflammatory activities of hesperidin will be able to encourage the clinical trials for neurodegenerative diseases. Further studies are necessary to develop medicine and/or medical supply for effective absorption of hesperidin, which owns various functions.

 XML Download
XML Download