

 PDF
PDF Citation
Citation Print
Print



Abbreviations
AIDP
acute inflammatory demyelinating polyneuropathy
APC
Ag-presenting cell
ASC
apoptosis-associated speck-like protein containing a caspase recruitment domain
BPI
bactericidal permeability increasing
CadF
cadherin-fibronectin binding protein
CAP
cationic antimicrobial peptide
Fn
fibronectin
GBS
Guillain-Barré syndrome
GI
gastrointestinal
GROα
growth related oncogene alpha
hBD
human β-defensins
IBS
Irritable bowel syndrome
IEC
intestinal epithelial cell
IP-10
IFN-γ-inducible protein 10
IRF-3
IFN regulatory factor 3
LOS
lipooligosaccharides
LR
lectin receptor
LTB4
leukotriene B4
MAMP
microbe-associated molecular pattern
MAP3K
mitogen activated protein kinase kinase kinase
NLRP3
NOD-like receptors with pyrin domain-containing 3
NOD
nucleotide-binding oligomerisation domain
OMV
outer membrane vesicle
PAMP
pathogen-associated molecular pattern
PRR
pathogen recognition receptor
RNS
reactive nitrogen species
Sn
sialoadhesin
Tc
cytotoxic T
TIR
Toll/IL-1 receptor
TRAF6
TNF receptor-associated factor 6
TRAM
Toll/IL-1 receptor-domain-containing adapter-inducing IFN-β-related adaptor molecule
TRIF
Toll/IL-1 receptor-domain-containing adapter-inducing IFN-β
INTRODUCTION
Campylobacter is commensal in poultry, but pathogenic in humans (12). The annual estimated number for Campylobacter infection cases is 96 million worldwide (2). Campylobacter is not harmless even for chickens as it stimulates the innate and adaptive immune responses in almost all types of chicken breeds. However, the extent of harm posed by Campylobacter may vary among different breeds of chickens (3). The differential susceptibility to Campylobacter across different breeds of chicken can be associated to the variation in their diet and gut microbiota composition (45). Chickens become colonised with Campylobacter at the age of 2–3 wk. Due to a lack of a fully developed adaptive immune system at this age, maternal Abs, already passed from hens to chicks, provide protection against Campylobacter (67). Maternal Abs against the Campylobacter flagellar proteins, outer membrane proteins and lipooligosaccharides (LOS) were observed in new-born chicks (7). After developing an adaptive immune system at the age of 6–7 wk, chickens produce Abs against Campylobacter cellular components, such as outer membrane proteins and flagellum (78). However, circulation of maternal Abs as well as development of adaptive immune B-cells play a limited role in the clearance of Campylobacter cells from the chicken intestines (69). It is proposed that Campylobacter avoids rapid clearance in the chicken intestine due to the adaptation to a novel colonisation mechanism, where it continues short-term invasion of chicken intestinal cells followed by escape from these cells (10). Clearance of Campylobacter from the chicken intestines may take many weeks, causing its persistence in chickens beyond slaughter age. Therefore, a poultry flock contaminated with Campylobacter is considered as a major source of Campylobacter transmission to humans (911).
The gastrointestinal (GI) tract in humans structurally comprises of four cell layers: the mucosa, submucosa, muscularis externa, and serosa. The mucosa is the innermost layer which is further divided into the epithelium, lamina propria, and muscularis mucosae. The mucosal epithelium of the small intestine comprises of a single layer of intestinal epithelial cells (IECs), mucus secreting goblet cells, paneth cells, and M-cells. The epithelium forms villi or crypts (finger-like projections) which are covered by a highly viscous mucus layer (12). The Campylobacter adherence to the mucus layer as well as its invasion into the IECs and lamina propria occurs mainly due to its mucins (that are glycosylated proteins in the mucus layer) degrading enzymes, flagella, and adhesins (which primarily include Jejuni lipoprotein A, fibronectin [Fn], Fn-binding protein FlpA, cadherin-Fn binding protein [CadF], cell binding factor 1, and cell-surface glycans) (1314151617). Following invasion into the host cells, Campylobacter reside into the LAMP-1 (late endosome marker) expressing Campylobacter-containing vacuoles (1819). The adherence, invasion and intracellular survival facilitate the cellular translocation (endocytosis and exocytosis) of Campylobacter within the epithelial cells. Campylobacter translocation is coupled with the release of toxins in order to trigger the ion instability, cell apoptosis, and pore formation in host cells (2021). The Campylobacter translocation and its potential to secrete toxins into host cells are the two main features of Campylobacter pathophysiology which enhance the inflammation and fluid secretion in intestinal cells and contribute to the development of infection in humans (16202122). Campylobacter infection in humans is characterised by an acute, self-limiting gastroenteritis which lasts for 5 to 7 days. The abdominal pain, watery or bloody diarrhoea, headache, fever, chills, and dysentery together with stools containing leukocytes and erythrocytes are the major signs of severe Campylobacter infection (23242526). Campylobacter infection can improve the progression of various persistent diseases including Guillain-Barré syndrome (GBS), Miller Fisher syndrome, Reiter's arthritis, and Irritable bowel syndrome (IBS) in humans. The LOS-outer core structures of Campylobacter are variable and mimic the structures of human neuronal gangliosides and for this reason, Abs produced against the LOS structural epitopes do not only bind to LOS structures, but also to human gangliosides. The cross-reactivity or non-specific binding of anti-LOS Abs with human neuronal gangliosides forms the basis of neural diseases specifically GBS development in humans (Fig. 1) (27282930).
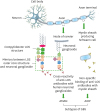
Figure 1
An illustration of interactions of anti-LOS Abs with human neuronal cells, likely to cause GBS in humans post Campylobacter infection.
Cross-reactivity of anti-LOS Abs occurs due to the mimicry between Campylobacter cell surface LOS core structures and human neuronal (node of ranvier) gangliosides and it develops a GBS subtype, AMAN. In some cases, anti-LOS Abs non-specifically bind to the Schwann cells to develop another type of GBS known as AIDP.
AIDP, acute inflammatory demyelinating polyneuropathy; AMAN, acute motor axonal neuropathy.
The scope of this review is to summarise available data on human immune responses involved in the defense against Campylobacter and how Campylobacter cells oppose these immune responses during infection. This review explains the role of human immune system in the elimination of Campylobacter infection by highlighting: i) induction of signalling pathways in intestinal mucosa for pathogen recognition; ii) influx of professional phagocytes into intestinal submucosa for bacterial clearance; iii) activation of adaptive immunity for persistent infection; iv) activation of serum proteins for persistent infection; and v) networking between human cells during Campylobacter infection.
INDUCTION OF SIGNALLING PATHWAYS IN INTESTINAL MUCOSA FOR PATHOGEN RECOGNITION
Campylobacter cells colonise the crypts in high numbers, rather than the intestinal lumen, due to the low concentration of oxygen and nutrients availability for maximal growth in crypts (1731). The pathogen recognition receptors (PRRs) of IECs become activated to recognise the pathogen-associated molecular patterns (PAMPs) when pathogens develop interaction with IECs extracellularly or survive intracellularly after invasion. The signalling pathways downstream the PRRs produce inflammatory and anti-inflammatory cytokines to regulate the immune responses during infection (323334). In addition to the activation of PRRs of IECs, increase in the number of mast cells and eosinophils has also been observed in lamina propria because they sense the Campylobacter cells as danger signals (3536).
Stimulation of cell-surface receptors in IECs
TLRs are the cell membrane bound PRRs which recognise the PAMPs during Campylobacter infection and consequently, induce the secretion of different interleukins and chemokines from IECs (32). Different cellular constituents of Campylobacter such as lipoproteins (bind TLR-1/2/6), LOS (bind TLR4), DNA (bind TLR9), capsule, cell wall polysaccharides, flagella, and toxins can bind to TLRs in human IECs to activate them (373839404142434445). The signalling pathway activates due to PAMPs binding to TLRs which recruits an adaptor protein, MyD88, to interact with IL-1 receptor-associated kinase complex and TNF receptor-associated factor 6 (TRAF6) (4647). This interaction induces the mitogen activated protein kinase kinase kinase (MAP3K) which further stimulates the MAPK including ERK and p38 induction. These MAPK translocate to the nucleus, act as transcription factors, and regulate the transcription of NF-κB as well as synthesis of pro-inflammatory cytokines (IL-8 and TNFα) in IECs (374148495051). The IECs TLRs (mainly TLR2) also involve the MyD88-independent signalling where they interact with adaptor proteins, Toll/IL-1 receptor (TIR)-domain-containing adapter-inducing interferon-β (TRIF) and TRIF-related adaptor molecule (TRAM), and activate the IFN regulatory factor 3 (IRF-3) (525354). IRF-3 accumulates in the nuclei and stimulates the synthesis of ILs (e.g. IL-1α and IL-6) via coordination with NF-κB (3252). During infection with Campylobacter, IECs also secrete chemokines including growth related oncogene alpha (GROα), MIP-1, MCP-1, and IFN-γ-inducible protein 10 (IP-10). This is also known to occur by the regulation of NF-κB transcription (555657); however, the role of specific TLRs in the induction of chemokines in IECs following infection with Campylobacter has not been yet investigated. Campylobacter jejuni (C. jejuni) can activate NF-κB independently of TLR signalling pathway (58), which might be linked to the chemokines production in Campylobacter infected IECs.
Stimulation of intracellular receptors in IECs
Campylobacter maintain their survival within the vacuoles (distinct from canonical endocytic vacuoles) inside of IECs (19). Direct invasion and subsequent intracellular survival of Campylobacter into IECs as well as the release of outer membrane vesicles (OMV) from extracellular Campylobacter cells cause delivery of the cellular constituents of Campylobacter into cytosol of host cells (185960). The nucleotide-binding oligomerisation domain (NOD) are the intracellular PRRs in IECs which directly recognise the microbe-associated molecular patterns (MAMPs; Campylobacter toxins, flagella, muramyl dipeptides, and adhesins) of Campylobacter into cytosol and induce the release of antimicrobial peptides, particularly human β-defensins (hBD)-2 (61626364). The hBD-2 are bactericidal as they disrupt the Campylobacter cell wall integrity (65). In addition, NOD1 binding to Campylobacter MAMPs also promotes the secretion of IL-8 from human IECs (373861) by activating the MAPK (1951). The OMV enclosed LOS, toxins, and N-linked glycoproteins, after their delivery into host cells, can also activate MAPK, particularly p38, to induce the secretion of IL-8, IL-6, TNF-α, and hBD-3 from IECs (59). IL-8 subsequent to its release recruits the innate immune cells including neutrophils, macrophages, and dendritic cells at the site of infection (343741485066). Inflammasome, a type of NOD-like receptor, is known to induce the pro-inflammatory cytokines (IL-1β and IL-18) in human cells (67). A recent study has demonstrated that inflammasome play a significant role in clearing the intracellular Campylobacter cells from human IECs (68), however, inflammasomes dependent specific mechanisms or signalling pathways involved in bacterial clearance are yet unknown.
INFLUX OF PROFESSIONAL PHAGOCYTES INTO SUBMUCOSA FOR BACTERIAL CLEARANCE
The intestinal crypts and IECs undergo the inflammation and severe damage during Campylobacter infection. In response to the inflammation and damage, IECs secrete cytokines and chemokines, which further recruit the professional mononuclear phagocytes including macrophages, neutrophils, NK cells and dendritic cells into the intestinal submucosa. These immune cells infiltrate the submucosal lining, interact with each other through cytokines, and help in clearing the bacterial cells from epithelium (23256669). A complex network of cytokines, linking IECs-to-immune cells and immune cells-to-each other, is presented in Fig. 2 together with key immune responses important for defense against Campylobacter.
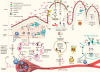
Figure 2
A representation of key immune responses, important for defense in humans against Campylobacter infection. The MyD88 dependent and independent signalling pathways downstream the IECs TLRs are involved in the synthesis of interleukins and chemokines via coordination with NF-κB. The cytosolic receptors (NOD1) and OMV also contribute to the activation of NF-κB for the synthesis of β-defensins. IECs-derived cytokines recruit neutrophils, macrophages, NK cells, and dendritic cells into the intestinal submucosa. The stimulation of MyD88 dependent and independent signalling downstream the TLRs in dendritic cells lead to the secretion of different cytokines, which further activate B-cells in a T-cell independent manner. The activation of MyD88 dependent signalling downstream the cell-surface receptors (TLRs and Sn) and stimulation of cytosolic receptors (NLRP3 inflammasomes) occur in macrophages during Campylobacter infection in order to produce inflammatory cytokines. The mature dendritic cells present Ags by MHC type molecules and secrete interleukins (IL-10 and IL-12) to increase the proliferation of naïve T-cells into Tc and Th cells. The interaction of dendritic expressed siglec-7 with Campylobacter LOS promote the differentiation of Th cells into Th1 and Th2. T-cells activate B-cells to produce Abs specific to the Campylobacter cell constituents. Abs from B-cells, IL-6 from macrophages and high level of acute-phase proteins in serum contribute to the activation of complement systems. Cells are associated to each other by a complex network of cytokines to regulate the immune responses. Cell-to-cell association is demonstrated by large brown arrows.
ASC, apoptosis-associated speck-like protein containing a caspase recruitment domain; BPI, bactericidal permeability increasing; GROα, growth related oncogene alpha; IP-10, IFN-γ-inducible protein 10; MAP2K, mitogen-activated protein kinase kinases; MAP3K, mitogen activated protein kinase kinase kinase; RNS, reactive nitrogen species; Sn, sialoadhesin; TIRAP, Toll/IL-1 receptor domain-containing adapter protein.
Neutrophils
The IECs-derived IL-8 is a neutrophil chemotactic factor, which induces the influx of neutrophils into the intestinal submucosa and infiltration of these neutrophils to the site of inflammation. In Campylobacter, methionyl-tRNA formyltransferase (encoded by fmt gene) is involved in the production of n-formyl peptides. These Campylobacter n-formyl peptides along with a host cell enzyme, 12-lipoxygenase, direct the migration of neutrophils from the basolateral to apical surface of the epithelium (66). A recent study demonstrates that the secretion of IL-10 downstream of the PI3Kγ signalling pathway in IECs can also play an important role in the infiltration of neutrophils into intestinal crypts and submucosa (70). The accumulated neutrophils present CD11b (cell-surface markers), which is a sign of initiation of phagocytosis process (71). They phagocytose complement-opsonised bacteria more efficiently rather than the non-opsonised bacteria (727374). They release toxic radicals or ROS (superoxide and hydrogen peroxide) for oxidative killing of bacteria and various inflammatory molecules including cationic antimicrobial peptides (CAPs), defensins, cathelicidins, bactericidal permeability increasing (BPI) protein (55 kDa), and leukotriene B4 (LTB4) for non-oxidative killing of bacteria (717576). The production of ROS and inflammatory molecules from neutrophils damage IECs structurally and contribute to the abscesses formation and loss of function in crypts (7177). This indicates that neutrophils are critical for the development of diarrhoea during Campylobacter infection. It is suggested that the non-invasion strains of Campylobacter can induce less concentration of IL-8 to further produce neutrophils and LTB4 in lesser amounts, which can lead to the development of non-inflammatory diarrhoea in humans rather than the inflammatory diarrhoea (78).
Monocytes/macrophages
Human monocytes with a range of cell-surface markers (CD14, CD11a) can be found in the intestinal mucosa following infection with Campylobacter for phagocytosis of Campylobacter cells (5079). Human macrophages (differentiated monocytes) are more important than the complement system for Campylobacter infection (79) and have ability to phagocytose the whole-bacterial cells, unlikely to neutrophils (727374). Monocytes undergo apoptosis following infection with Campylobacter, however, macrophages rapidly kill Campylobacter cells subsequent to their internalisation (19738081). Campylobacter cells in coccal or degenerative form can be observed in macrophages after 4–8 hours of infection (72). Human macrophages generally possess cell membrane bound receptors (TLR and lectin receptors [LRs]) and cytosolic receptors (inflammasomes) (334482), which become activated to recognise the PAMPs when Campylobacter cell develops interaction with a macrophage or survive intramacrophage subsequent to the phagocytosis (3383). Campylobacter LOS, cell wall polysaccharides, lipoproteins, and N-linked glycosylated proteins have been reported as ligands of macrophage TLR4, sialoadhesin (Sn; a type of LRs), and galactose-type lectin receptors (another type of LRs) (144481828485). These cellular components detached from killed Campylobacter cells as well as viable cells of Campylobacter can bind to macrophage (specifically M1) receptors and induce TLR-MyD88 dependent signalling pathway in order to secrete the pro-inflammatory cytokines (IL-1α, TNF-α, pro-IL-1β, and IL-6) and proteins (such as NOD-like receptors with pyrin domain-containing 3 [NLRP3] proteins). M1-derived IL-6 helps in the activation of complement system while other cytokines recruit more innate immune cells to the site of inflammation. The NLRP3 proteins, after release into cytosol, combine with apoptosis-associated speck-like protein containing a caspase recruitment domain (ASC) and pro-caspase1 to assemble NLRP3-inflammasomes (protein complexes). Upon a second signal, NLRP3-inflammasomes activate caspase-1 to catalyse the 31 kDa pro-IL-1β (already located into cytosol) into mature, biologically functional 17 kDa IL-1β. The cleaved IL-1β is then secreted outside the cell to mediate the inflammatory responses (143344508185). Further, IFN-γ (which is released from IECs and T-cells during actual infection) leads to the production of nitric oxide synthase 2 and reactive nitrogen species (RNS) in M1 macrophages, which are potent and effective towards killing of Campylobacter cells (348687). During Helicobacter pylori infection, M2 macrophages and cytokines (IL-10, IL-12, and TGFβ) production from these M2 cells can be stimulated by T-cells derived anti-inflammatory cytokines (such as IL-4, IL-10, and IL-13) in order to suppress further activation of immune cells and inflammation (8889). In the same way, M2 macrophage polarisation can play its role in immunity and inflammation suppression during Campylobacter infection, however, it needs to be confirmed by further research.
NK cells
NK cells could be a source of cytokines, such as IFN-γ, but its association to Campylobacter cells has not been yet investigated. Siglec 7 present on the cell surface of NK cells could be potential receptors involved in the development of pathogen-host cell interaction (74).
Dendritic cells
Dendritic cells reside under IECs and can extend themselves between IECs to sample the lumen, therefore, they are well positioned to interact with Campylobacter cells present inside of intestinal lumen and submucosa (9091). Dendritic cells form an early line of defense within the submucosa against the invasive Campylobacter strains, as well as, in the intestinal lumen against the non-invasive Campylobacter strains (124092). Dendritic cells readily internalise the Campylobacter cells, express cell surface co-stimulatory molecules (CD14, CD40, CD80, and CD86), and become mature (9394). TLRs in mature dendritic cells (Ag-presenting cells; APCs) can become activated as a consequence of their internalisation of whole-bacterial cell or their interaction with bacterial cellular components. The MyD88-dependent signalling pathway downstream the TLR2 and TLR4 produce different pro-inflammatory cytokines including IL-1, IL-6, IL-8, IL-10, IL-12, IFN-γ, and TNF-α, while, the MyD88-independent signalling via TLR4-TRIF axis contributes to the production of IFN-β in mature dendritic cells (408795). It is proposed that C. jejuni cell-surface sialylated LOS structures intensify the production of IFN-β and TNF-α in dendritic cells which further contribute to the proliferation of human mucosal B-cells in a T-cell independent manner. It may link the C. jejuni sialylated LOS structures with the initiation of B-cells mediated autoimmunity in GBS (9394). Other cell membrane bound receptors, siglec-7, of mature dendritic cells can also interact with Campylobacter LOS to increase the cytokines production and uptake of Campylobacter cells into dendritic cells (74), but the mechanisms involved downstream of the siglec-7 activation in mature dendritic cells are not yet known.
ACTIVATION OF ADAPTIVE IMMUNITY FOR PERSISTENT INFECTION
The adaptive immunity develops typically to eradicate the persistent Campylobacter infection as well as to reduce the post-infection severe complications (96). It is supported by the presence of high amount of intraepithelial T-lymphocytes in patients with post-dysenteric IBS, acute inflammatory demyelinating polyneuropathy (AIDP), and colorectal cancer (979899). The APCs after pathogen recognition migrate to the lymphoid nodules (Peyer's patches) where they present Ags via cell-surface molecules (MHC-I and MHC-II) to trigger the polarisation of naïve T-cells to CD8+ cytotoxic T (Tc) cells and CD4+ Th cells. Further, Campylobacter LOS structures bind with cell-surface siglec-7 receptors of APCs to mediate the differentiation of Th cells into the Th1 and Th2 cells (4087). It has been reported that APC-expressed siglec-7 receptors interaction with α2, 8-linked sialylated LOS induces the Th1 polarisation, whereas, its interaction with α2, 3-linked sialic acid induces a Th2 development (87). Moreover, cytokines (IL-12 and IL-10) from APCs also stimulate the proliferation of Th1 cells and secretion of IFN-γ, TNF-α, IL-22, and IL-17 from these T-cells. Th1 cells activate more Tc cells and macrophages to enhance immunity against the invading or intracellular microbes, whereas, Th2 mediate class switching in B-cells to enhance immunity against the extracellular microbes. The cytokines from dendritic cells do not only induce the activation and proliferation of B-cells in a T-cell dependent manner, but also in T-cell independent manner. Hence, dendritic cells act as a bridge between innate and adaptive immune systems (4087100101102). Apart from Th1 and Th2 cells, frequency of other types of CD4+ Th cells including Th17, Th22, and Treg and their associated cytokines (IL-17, IL-18, IL-22, and IL-23) has also been observed elevated in patients serum following infection with Campylobacter (103104). Subsequently, B-cells after activation produce Abs against the Campylobacter toxins, flagella, LOS, CadF, and major outer membrane proteins, and secrete them in human serum (729105106). In the acute phase of infection (7 days post-infection), the level of serum Abs, IgA, and IgM, increase in serum (107). In the convalescent phase of infection (1 wk to 2 months), IgG also begins to circulate in the blood (108109). These serum Abs are detectable in the serum and faeces of Campylobacter infected patients (110). IgA in up to 20 days and IgM in 2 months attain their normal levels back. In contrast, IgG present in serum as well as serum IgG expelled into saliva, remain extant inside the host for long time period (1 year) and provide protection against subsequent Campylobacter infection (108109).
The LOS-outer core structures (GM1, GM2, GM3, GD3, or GD1-like) present on the cell-surface of Campylobacter mimic the GM1, GM2, GM3, GD3, and GD1 containing human neuronal gangliosides. Campylobacter possess phase variation in LOS biosynthesis genes and therefore, can switch the LOS-outer core structures from one form to other (GM1⇔GM2; GM2⇔GM3; GD1⇔GD3). The LOS-outer core structures' mimicry with human neuronal gangliosides and their switching ability help Campylobacter to escape from the host immune system (30111112). It has been identified that Campylobacter toxins arrest human T-cells in the G2 phase of cell cycle and halt their development (113). The representation of mimics of human neuronal gangliosides on cell-surface, ability to vary these mimics, and toxins mediated inhibition of T-cells indicate that Campylobacter has evolved strategies to escape from the host adaptive immunity.
ACTIVATION OF SERUM PROTEINS FOR PERSISTENT INFECTION
Bacteria opsonised by the professional phagocytes (macrophages, neutrophils, dendritic cells) can enter into blood stream and can be circulated back to neutrophils as neutrophils mediate killing of opsonised bacteria more efficiently than the non-opsonised ones. Defense system in humans involving innate and adaptive immune responses limit the infection to the site of inflammation and does not allow live microbes to enter into bloodstream (7273). However, during Campylobacter infection, bacteremia can be developed in those patients which are immunocompromised or have persistent post-infection complications. The acute phase proteins particularly C-reactive proteins have been found elevated in GBS patients and patients with weak immune system (such as Campylobacter infected children), which are further likely to activate the complement system (114115). Serum proteins involved in both classical and alternative complement pathways are bactericidal and can facilitate direct killing of Campylobacter. Campylobacter capsule can provide protection to Campylobacter against killing mediated by serum or complement proteins (76116117118). Human C3b proteins were found unable to bind to the encapsulated Campylobacter fetus previously, supporting the role of capsule S-layer proteins in the development of interaction with human serum proteins (119).
NETWORKING BETWEEN HUMAN CELLS DURING CAMPYLOBACTER INFECTION
During Campylobacter infection, networking between IECs and immune cells as well as among different immune cells occurs with the help of cytokines (Fig. 2). In Fig. 2, it has been demonstrated that: i) the Campylobacter infected IECs release IL-8 and IL-10 to recruit neutrophils into lamina propria and submucosa; ii) different cytokines including IL-10, IL-12, TNF-α, and IFN-β from dendritic cells activate B-cells in both T-cell dependent and independent manners; iii) the IFN-γ secretion from Th1 and Tc cells stimulate more macrophages during infection; and iv) the production of IL-6 and Abs respectively from macrophages and B-cells as well as a high level of acute-phase proteins in serum contribute to the activation of complement system (40667072868794105).
The GI tract in humans express hBD-1 constitutively, while, the production of other defensins (hBD-2 and hBD-3) involves cell receptors mediated signalling pathways, NF-κB transcription, and cytokines secretion (65). During Campylobacter infection, hBD-1 expression remains unchanged. However, expression of other defensins increases due to stimulation of IECs membrane bound receptors (TLR) and intracellular receptors (NOD1) following their interaction with Campylobacter cells or cellular constituents (6165). The level of β-defensins in IECs further rises through networking between different immune cells (dendritic cells, CD14+ macrophages, Tc, and NK cells) and IECs (Fig. 3) (100101).
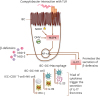
Figure 3
Networking between human IECs and innate immune cells for maximal production of β-defensins in IECs.
The presence of Campylobacter OMV inside of an IEC, activation of intracellular NOD-like receptors (NOD1), and an interaction between the host IEC TLR and a Campylobacter cell, all lead to the activation of MAPK. The IECs and immune cells collaborate to secrete a triad of cytokines (IFN-γ, IL-22, and IL-23), which further elevates the expression of IL-17 in IECs. The activated MAPK and a high level of IL-17, both stimulate the production of β-defensins in IECs by the regulation of NF-κB transcription.
GAPS IN KNOWLEDGE FOR FUTURE RESEARCH
This review identifies that substantial gaps are present in that knowledge which relates the human protective immunity to Campylobacter infection. These knowledge gaps have not so far been investigated and can be filled by future research. The association of IECs-derived cathelicidins and bactericidal permeability-increasing proteins to Campylobacter infection has not been established to date (12). The IECs cell receptors and signalling pathways, important for the induction of chemokines during Campylobacter infection, have not been yet identified. The inflammasomes activation in IECs in response to Campylobacter infection as well as their functions have been reported previously (68), but further investigations are required to identify the stimulatory factors of inflammasomes and mechanisms involved subsequent to their stimulation. A link between the Campylobacter OMV and inflammasome induction inside of IECs might be present and can be focused in future studies. Similarly, Campylobacter OMV might also have implications for other NOD-like intracellular receptors (e.g. NOD1). In addition, the activation of siglec-7 receptors in dendritic cell, acute phase proteins in serum, and T-cell subsets in lymphoid nodules during Campylobacter infection and their related host responses are yet to be explored in detail. Moreover, a possible connection present between many acute phase proteins (alpha 1-antitrypsin, mannose-binding lactin, and serum amuloid A) and Campylobacter infection has never been investigated. An outer membrane protein of C. jejuni, CadF, facilitates interaction between the host cell fibronectin and Campylobacter cells (15), which might be a target of serum amyloid A (12). This prediction requires verification with further research.
 XML Download
XML Download