

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Although acute flexor tendon injuries are relatively uncommon, restoration of hand function following such trauma has been challenging.123 Adhesions may form between the repaired tendons and surrounding tendon sheath, especially in the tight fibro-osseous spaces of zone II injuries, despite meticulous surgical techniques and aggressive postoperative therapy.45 Several innovative approaches to prevent adhesion formation after tendon repair have been proposed, including anti-inflammatory agents, such as steroids and non-steroidal anti-inflammatory drugs;67 natural polymers, such as hyaluronic acid,8910 collagen solution,11 and amniotic fluid;11 anti-metabolites, such as 5-fluorouracil;1213 and barrier methods ,such as polytetrafluoroethylene membrane and porcine gelatin.1415 However, these anti-adhesion therapies remain unsuccessful and have not progressed beyond preclinical studies.
It has been shown that the transforming growth factor-beta (TGF-β) pathway is involved in the formation of adhesions.16 Studies on TGF-β pathway inhibition have reported an improvement in the range of digital motion as well as alteration of types I, III, and V collagen production.1718 In vitro studies using the natural TGF-β inhibitors decorin and mannose 6-phosphate (M6P) have reported a reduction in TGF-β induced collagen production, especially type I collagen.19 However, other studies have reported that TGF-β pathway inhibition is associated with a decrease in mechanical strength,2021 and a clinical trial on M6P did not demonstrate any beneficial effect on finger movements.22
It has been reported that activin A in fibroblasts, which is induced by TGF-β1, stimulates inflammation and fibrosis in many different models, including inflammatory bowel disease, rheumatoid arthritis, and wound healing, and following burn injuries.2324 A key regulator of activin is follistatin (FST), a glycoprotein that binds activin. This binding is virtually irreversible. The resulting complex targets a lysosomal degradation pathway.25 FST can block the fibrogenic actions of TGF-β despite its inability to bind TGF-β1, suggesting that TGF-β1 exerts fibrogenic actions through its stimulation of activin A.24 In addition, as FST binds to activin A, it attenuates the capacity of activin A to stimulate fibrosis, as demonstrated in hepatic fibrosis,26 pulmonary fibrosis,27 and radiation-induced fibrosis.28
In this study, we hypothesized that the activin/FST pathway offers a novel biological target for preventing the formation of tendon adhesions. We used an adenovirus system to transfer a follistatin gene construct into fibroblasts isolated from scar tissue around the repaired flexor tendons of patients who underwent tenolysis after repair of zone II tendon. The goal of this experiment was to investigate the effect of this gene on inhibition of fibrosis.
MATERIALS AND METHODS
The Human Subjects Institutional Review Board of Yonsei University Health System, Severance Hospital (4-2016-0174) approved all of the experimental protocols in this study.
Study design
To test the anti-fibrotic effect of adenovirus-FST construct (Ad-FST) on fibroblasts in vitro, we utilized cells from the scar tissue of patients who underwent tenolysis following repair of zone II flexor tendons. Fibroblasts were activated by TGF-β1 (2 ng/mL), exposed to Ad-FST as a therapeutic gene and adenovirus-LacZ construct as a marker gene, and cultured for 24 hours. The second set of fibroblast cultures that were not exposed to Ad-FST or Ad-LacZ served as the control. The mRNA expression levels of collagen types I, III, IV, V, and XI, as well as MMP-1, -3, -8, -9, and -13 were analyzed by reverse-transcription polymerase chain reaction (RT-PCR). In addition, protein expression levels of fibronectin, PAI-1, TRPV4, MMP-1, -2, -13, TIMP-1, -2, -4, and alpha-smooth muscle actin (α-SMA) were estimated using Western blot analysis.
Isolation and cell culture
Scar tissues around the repaired zone II flexors were harvested from seven patients, in whom adhesions had formed between the tendon and its sheath during tenolysis. Minced tissues were digested at 37℃ under gentle agitation with type IV collagenase (250 unit/mL; Sigma, St. Louis, MO, USA). Cells were placed in culture plates (TPP, Trasadingen, Switzerland) at 4×105 cells/mL. Primary cultures were sustained for 2–3 weeks in Dulbecco's Modified Eagle Medium (DMEM; Life Technologies Corporation, Carlsbad, CA, USA) containing 10% fetal bovine serum, 1% v/v penicillin, streptomycin, and nystatin (Life Technologies Corporation) in a 37℃ incubator with 5% CO2 and humidity. The isolated primary cells at passage 1 were subcultured at a 1:3 ratio when they were 80–90% confluent, up to P3, which were stored at −80℃ until being used.
Follistatin constructs and transfections
For this study, Ad-FST expressing the human follistatin gene was prepared, and Ad-LacZ expressing the LacZ gene was used as a viral control. Each recombinant adenoviral vector originated from replication-deficient adenovirus type 5 lacking E1 and E3 regions (Fig. 1). FST gene was cloned into the E1 region under the control of human cytomegalovirus early promoter. Recombinant virus was grown in transformed human embryonic kidney 293 (HEK293) cells and purified with Vivapure AdenoPACK 100 (Sartorius Stedim Biotech, Goettingen, Germany). Titers were determined by optical density at 260 nm. At confluence, fibroblastic cells were rinsed with Dulbecco's phosphate buffered saline (PBS; Life Technologies Corporation), and then exposed to DMEM containing one dose of Ad-LacZ or Ad-FST with a viral infection multiplicity of 80.
RT-PCR analysis for matrix metalloproteinases and collagens
Total RNA was isolated from the fibroblasts transfected with Ad-FST for 24 hours using QIAGEN RNeasy mini kit (QIAGEN, Germantown, MD, USA). In addition, cDNA was prepared using QuantiTect Reverse Transcription Kit (QIAGEN). Collagen and MMP genes were amplified, and relative expressions were normalized to the average beta-actin levels. Resulting data were analyzed using ImageJ software version 1.6 (National Institutes of Health, Bethesda, MD, USA).
Protein extraction and Western blot analysis
Fibroblasts at a density of 2×105 cells per well were transfected with Ad-FST for 24 hours and lysed in RIPA lysis buffer (ATTO Corp., Tokyo, Japan) containing protease inhibitor (Pierce Mini Tablets, IL, USA). Meanwhile, the culture medium was collected to measure FST protein expression by Ad-FST. Lysates and culture medium were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis, and proteins were then transferred to polyvinylidene difluoride membranes (Merck Millipore Ltd., Darmstadt, Germany) using a transfer system (Mini Trans-Blot Cell and systems, Bio-Rad, Hercules, CA, USA). The blots were incubated with specific antibodies against MMP-1, MMP-13, TIMP-1, TIMP-2, TIMP-4, fibronectin, PAI-1, TRPV4, and α-SMA (Abcam, Cambridge, UK). After reacting with secondary antibodies, immunoreactive bands were visualized by a Western blot detection system (EzWest-Lumi Plus, ATTO Corp., Japan). To verify the amounts of loaded proteins, blots were stripped of bound antibodies and reprobed using antibodies to actin (Abcam).
Immunocytochemistry
The cells were transfected with Ad-FST for 24 hours at a density of 2×105 cells per well in culture medium, and fixed in 4% formaldehyde. The cells were then permeabilized in 0.1% Triton X-100 (SIGMA, St. Louis, MO, USA) for 20 minutes, and incubated in PBS with 1% bovine serum albumin for 1 hour. The cells were incubated with antibodies overnight at 4℃. The secondary antibodies (stained in red and green) were used at a 1:1000 dilution for 1 hour. DAPI was used to stain the cell nuclei (blue).
Statistical analyses
All data are expressed as the mean±standard deviation compiled from three independent experiments of separate cultures isolated from seven donors. A two-tailed Student's t-test was used to compare the means of the two groups. A p value<0.05 was considered statistically significant.
RESULTS
FST protein expression from Ad-FST transfected fibroblasts
Fibroblasts from the scar tissue of patients with adhesions around a repaired zone II flexor tendon that were transfected with Ad-FST showed increased staining of FST protein on confocal images compared to fibroblasts transfected with a control (Fig. 1), indicating that FST gene construct was efficiently transduced into the fibroblasts.
Expression of collagen type I, MMP-1, and MMP-2 mRNAs
Fibroblasts transfected with Ad-FST exhibited a significant (25%) decrease in collagen type I mRNA expression at 24 hours compared to Ad-LacZ controls (TGF-β1+) (p=0.003) (Fig. 2). Fibroblasts transfected with Ad-FST also exhibited significant decrease in MMP-1 and MMP-2 mRNA expressions (31% and 59%, retrospectively) at 24 hours compared to Ad-LacZ controls (TGF-β1+) (Fig. 3). The expression of α-SMA mRNA also was significantly decreased (23%) in fibroblasts transfected with Ad-FST at 24 hours compared to Ad-LacZ controls (TGF-β1+) (p=0.001) (Fig. 4). The effect of TGF-β1 on these fibroblasts did not change when treated with a selective small molecule in hibitor, SB505124, of the canonical (i.e., Smad-dependent) TGF-β signaling pathway. These findings suggest that fibroblasts may be activated by TGF-β1 via the non-canonical TGF-β signaling pathway.
Expression of MMP-1, TIMP-1, fibronectin, PAI-1, TRPV4, and α-SMA
Fibroblasts transfected with Ad-FST exhibited significant decrease in MMP-1, TIMP-1, and fibronectin (24%, 23%, and 24%, retrospectively) at 24 hours compared to Ad-LacZ controls (TGF-β1+) (p<0.001, p=0.002, p<0.001, retrospectively) (Figs. 5 and 6). Fibroblasts transfected with Ad-FST also demonstrated significant inhibition of TRPV4, PAI-1, and α-SMA protein expressions (23%, 23%, and 28%, retrospectively) at 24 hours compared to Ad-LacZ controls (TGF-β1+) (p=0.002, p<0.001, p=0.002, retrospectively) (Fig. 7).
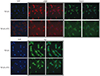
Immunocytochemistry analysis of α-SMA, desmin, and PAX7
Fibroblasts with Ad-FST exhibited decrease in α-SMA, desmin, and PAX7 expressions compared to Ad-LacZ controls (Fig. 8).
DISCUSSION
Following repair, tendons heal via both intrinsic and extrinsic pathways. Intrinsic healing involves the proliferation of tenocytes and the production of extracellular matrix (ECM).29 Extrinsic healing involves an invasion of cells from the surrounding sheath and synovium, which envelop the tendon structure and contribute to the formation of adhesions.29 Extrinsic healing dominates when the tendon or tendon sheath is injured, as the tendon has poor cell density and growth factor activity. Although several approaches have been proposed to prevent the formation of adhesions, there is limited evidence to prove that these approaches are successful, and none has been widely accepted for clinical use.30 Since tendon injury stimulates increased expression of TGF-β1 in the production of collagen types I and III, fibronectin, and glycosaminoglycans,31 blocking TGF-β1 has been reported to decrease tendon adhesion in animal tendon injury models. However, blocking TGF-β1 can also result in reduced tendon strength, as another role of TGF-β1 is to modulate immune responses and tissue repair.162032 Therefore, it is necessary to investigate methods that can selectively isolate and block the pro-fibrosis effects of TGF-β1.
FST, a systemically-expressed circulating glycoprotein, is the endogenous ligand-binding partner for activin A.33 It is well-recognized that FST can inhibit the activity of activin A, which is overexpressed in TGF-β1-induced fibrosis.2434 In the present study, the anti-fibrotic effect of FST was investigated via genetic modification of fibroblastic cells from patients with zone II flexor tendon adhesion after tendon repair. We found that fibroblastic cells from scar adhesion tissue transfected with Ad-FST showed decrease in expression levels of collagen type I mRNA, compared to fibroblasts transfected with Ad-LacZ controls. These findings indicate that FST gene inhibits the transcription of collagen type I. We also showed that the inserted FST gene decreased both transcription and translation of MMP-1 and TIMP-1. Tissue inhibitors of metalloproteinases (TIMPs) are pleiotropic extracellular proteins, and are recognized as endogenous regulators of matrix metalloproteinases (MMPs), which comprise a large family of extracellular enzymes with proteolytic activities that participate in cellular homeostasis, adaptation, and tissue remodeling.35 The findings of the present study suggest that FST restores the pathological imbalance of ECM turnover from zone II flexor tendon adhesions via the downregulation of MMP-1 and TIMP-1, which may contribute to the decrease in scar formation.
It is now accepted that myofibroblasts go through a precursor stage, in which large stress fibers are expressed. These large stress fibers are not observed in quiescent fibroblasts. These prominent bundles of microfilaments permit some contraction, pre-stressing, and remodeling of the surrounding ECM.36 At a later stage, fully differentiated myofibroblasts exhibit expression of α-SMA, which is usually an SM-specific cytoskeletal protein and is often used to define the myofibroblast phenotype.37 In the present study, the synthesis of fibronectin, a non-collagenous protein, as well as PAI-1 and α-SMA, were suppressed in fibroblasts transfected with Ad-FST compared to Ad-LacZ controls at protein level. The significant decrease in α-SMA expression suggests that FST may inhibit the transformation of fibroblasts to myofibroblasts. The decrease in transcription of collagen type I gene may also be associated with a decrease in the transformation of fibroblasts to myofibroblasts.
In the present study, the synthesis of TRPV4 was suppressed at protein level in fibroblasts transfected with Ad-FST compared to Ad-LacZ controls. It has been proposed that myofibroblasts contract via Ca2+-dependent mechanisms similar to those present in smooth muscle cells, with increased free Ca2+ regulating the phosphorylation of myosin light chain.38 The findings from our study suggest that FST may decrease the contraction of fibroblasts from tendon adhesion tissue by suppressing the translation of TRPV4.
In conclusion, FST may be a novel therapeutic strategy for preventing scar adhesions around repaired tendons by inhibiting fibroblasts from differentiating into myofibroblasts, as well as producing type I collagen and regulating ECM turnover via the downregulation of MMP-1 and TIMP-1. Due to the lack of knowledge regarding FST on tenocytes after tendon injury, further studies should be performed on how FST would affect tendon healing and strength when tenocytes are tested. FST may also decrease the contracture of the scar produced from myofibroblasts by inhibiting Ca2+-dependent contraction of the cells.







 XML Download
XML Download