

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
In humans, KRAS proto-oncogene is located at 12p12.1 and consists of 6 exons. It encodes guanosine triphosphatase (GTPase) KRAS isoform, which is a 21-kDa protein with 188 or 189 amino acids. KRAS protein is a member of the RAS superfamily of small GTPases [1]. A single amino acid substitution in KRAS is responsible for an activating mutation. It is known to be associated with human malignancies such as pancreas ductal carcinoma [2], colorectal carcinoma [3], lung adenocarcinoma [4], and various other human malignancies [56].
Little is known about the role of KRAS in breast cancer. Some of the previous studies have reported the potential role of KRAS mutation in endocrine resistance of luminal breast cancer [78]. Genome-wide screening results showing an association between KRAS status and breast cancer have been reported [91011]. Other studies have shown the association of KRAS 3′ untranslated region (UTR) variants with increased risk of breast cancer [121314]. The prognostic role of KRAS in breast cancer and the role of KRAS mRNA expression status per se in human malignancy remain unclear. Although many previous studies have determined the impact of the activating mutation of KRAS on human malignancies, little is known about the significance of other molecular statuses such as copy number alteration (CNA), methylation, mRNA expression, and protein expression. Therefore, the following 2 well-organized public databases were utilized for this study. The Cancer Genome Atlas (TCGA) Research Network has profiled a large number of human tumors including breast cancer to discover molecular aberrations at the DNA, RNA, protein, and epigenetic levels [15]. Regarding breast cancer, TCGA Research Network has previously reported the comprehensive molecular portraits of invasive ductal and invasive lobular carcinomas [910]. The Molecular Taxonomy of Breast Cancer International Consortium (METABRIC) has released genomic and transcriptomic architecture of 2,000 breast cancers [1617]. METABRIC database is believed to provide landscapes for understanding how somatic CNA affects gene expression and reveal novel subgroups that could be the target for future investigation in breast cancer.
The objective of this study was to investigate the prognostic role of KRAS mRNA expression in breast cancer using TCGA and METABRIC databases. Molecular regulation of KRAS was also investigated, including mRNA expression, methylation, CNA, and mutation.
METHODS
Data acquisition
TCGA and METABRIC databases were acquired from the following websites: TCGA (https://cancergenome.nih.gov/), Synapse (https://www.synapse.org/), and cBioPortal (http://www.cbioportal.org/). Currently, cBioPortal contains datasets of 233 cancer genomics studies including 14 breast cancer studies. TCGA dataset on breast invasive carcinoma (TCGA, Provisional) and METABRIC dataset on breast cancer (METABRIC, Nature 2012) were utilized for this study (access date: July 11, 2018). The Institutional Review Board of Seoul Metropolitan Government-Seoul National University Boramae Medical Center approved this study (approval number: 17-2018-23). The informed consent of this study was waived.
Clinicopathologic parameters
Patient age was defined as the age at the time of initial diagnosis of primary breast cancer. TNM categories and anatomic stage groups were described according to the breast cancer staging system of the American Joint Committee on Cancer. The status of estrogen receptor or progesterone receptor was described based on the result of immunohistochemical test for each receptor. Hormone receptor (HRc) status was defined according to the statuses of estrogen and progesterone receptors. Human epidermal growth factor receptor 2 (HER2) status was defined according to the results of immunohistochemical test and in situ hybridization assay. Regarding TCGA dataset, the breast cancer subtypes were classified according to the statuses of HRc and HER2. Regarding METABRIC dataset, the breast cancer subtypes were described according to the PAM50 classification.
Biological parameters
RNA sequencing (RNA seq) data were obtained from two different datasets: RNA seq median value by Illumina RNA seq version 2 RSEM and RNA seq z-score. RNA microarray (RNA mic) data were provided as median values of raw data by Agilent microarray analysis. Methylation data were provided as beta values of raw data. CNA linear (CNA_lin) data were provided as relative linear values for each gene by Affymetrix SNP 6. CNA non-linear (CNA non) data were provided as non-linear values by Genomic Identification of Significant Targets in Cancer (GISTIC) 2.0 with values of −2, −1, 0, 1, and 2, which represented homozygous deletion, hemizygous deletion, neutral or no change, gain, and high-level amplification, respectively. Mutation data were generated by whole exome sequencing and provided in a mutation annotation format. Biological data types and sample numbers utilized in this study are described in Supplementary Table 1. Biological parameters were classified into low or high based on the mean value of each parameter. RNA seq z-scores were classified into down-regulation, normal-regulation, and up-regulation with cut-off values of −2 and +2, respectively.
Statistical analyses
Two-sample t-test was used to determine the difference in expression levels of biological parameters while Pearson's χ2 test was used to determine the difference in clinicopathologic characteristics between groups. Pearson correlation coefficient (r) was used to evaluate the bivariate correlation between biological parameters and continuous variables. Spearman correlation coefficient (ρ) was used for biological parameters with ordinal variables. Survival analyses were carried out regarding overall survival (OS) and breast cancer-specific survival (BCSS). Kaplan-Meier estimation was used to analyze survival rates while log-rank test was used to determine the significance of differences between 2 survival curves. Cox-proportional hazards model was used for univariable and multivariable analyses. Hazard ratio (HR) was calculated using 95% confidence interval (CI). All statistical analyses were conducted using IBM SPSS Statistics version 20.0 (IBM Inc., Armonk, USA) and R software version 3.3.2 (R Foundation for Statistical Computing, Vienna, Austria). Statistical significance was considered when p-value was less than 0.05.
RESULTS
Clinicopathologic characteristics of study subjects according to expression level of KRAS mRNA
Total number of study subjects was 1,093 from the TCGA dataset and 1,904 from the METABRIC dataset. The mean follow-up period regarding OS was 40.9 ± 39.2 months (range, 0–283 months) from the TCGA dataset and 125.0 ± 76.3 months (range, 0–355 months) from the METABRIC dataset. The mean follow-up period regarding BCSS was 98.2 ± 60.4 months (range, 0–307 months) from the METABRIC dataset. Baseline clinicopathologic characteristics of study subjects are summarized in Table 1. In TCGA dataset, there was no significant difference in clinicopathologic factors except M category. In METABRIC dataset, proportions of subjects with positive estrogen receptor or positive progesterone receptor were higher in the group with low KRAS expression compared to those in the group with high KRAS expression while the group with high KRAS expression group showed higher proportion of subjects with positive HER2. Luminal A subtype was more prevalent in the group with low KRAS expression.
Table 1
Clinicopathologic characteristics according to expression level of KRAS mRNA in TCGA and METABRIC databases

Survival analyses according to expression level of KRAS mRNA
In TCGA dataset, the group with high KRAS mRNA expression showed worse OS compared to that shown by the group with low KRAS mRNA expression (p = 0.012, Figure 1A, cut-off value of RNA seq median value = 1,356.9). The group with up-regulated expression of KRAS mRNA also showed lower OS rate compared to that shown by the group with normal-regulated KRAS expression (p < 0.001, Figure 1B). In METABRIC dataset, the group with high KRAS mRNA expression showed worse survival rate than the group with low KRAS mRNA expression regarding OS (p < 0.001, Figure 1C) and BCSS (p = 0.001, Figure 1D). Similarly, the third and fourth quartiles (Q3 and Q4), according to the expression level of KRAS mRNA using RNA mic data, showed worse survival rates than the first or second quartiles (Q1 or Q2) regarding both OS and BCSS (Supplementary Figure 1). In METABRIC dataset, the group with high KRAS mRNA expression showed lower OS rate for luminal A subtype (p < 0.001, Figure 2A), but not for other subtypes (Figures 2B-D). Detailed survival rates are shown in Supplementary Tables 2 and 3. In TCGA dataset, the expression of KRAS mRNA was lower in luminal A subtype compared to that in any other subtypes (all p < 0.001, Supplementary Figure 2). There were no significant differences in clinicopathologic factors according to mRNA expression level for luminal A subtype (Supplementary Table 4).
Figure 1
Survival curves according to the expression level of KRAS mRNA using TCGA and METABRIC databases. OS curves according to RNA seq median value (A) and RNA seq z-score (B) in TCGA database. OS curves (C) and BCSS curves (D) according to RNA mic in METABRIC database.
OS = overall survival; TCGA = The Cancer Genome Atlas; METABRIC = Molecular Taxonomy of Breast Cancer International Consortium; RNA seq = RNA sequencing; BCSS = breast cancer-specific survival; RNA mic = RNA microarray.

Figure 2
OS curves according to the expression level of KRAS mRNA in each subtype of breast cancer using METABRIC database. OS curves in luminal A (A), luminal B (B), HER2 (C), and basal (D) subtypes.
OS = overall survival; METABRIC, Molecular Taxonomy of Breast Cancer International Consortium; HER2 = human epidermal growth factor receptor 2; RNA mic, RNA microarray.

Univariable and multivariable analyses
Expression level of KRAS mRNA was a significant factor in univariable analysis of both TCGA (HR, 1.501; 95% CI, 1.091–2.067; p = 0.013) and METABRIC (HR, 1.238; 95% CI, 1.100–1.394; p < 0.001) datasets. It was also an independent significant prognostic factor for breast cancer according to multivariable analysis of both TCGA (HR, 1.570; 95% CI, 1.026–2.403; p = 0.038) and METABRIC (HR, 1.254; 95% CI, 1.087–1.446; p = 0.002) datasets after adjusting for age, T category, N category, M category, estrogen receptor, progesterone receptor, and HER2 (Table 2).
Table 2
Univariable and multivariable analyses regarding OS using TCGA and METABRIC databases

OS = overall survival; TCGA = The Cancer Genome Atlas; METABRIC = Molecular Taxonomy of Breast Cancer International Consortium; HR = hazard ratio; CI = confidence interval; HER2, human epidermal growth factor receptor 2; HRc, hormone receptor; RNA mic, RNA microarray; RNA seq, RNA sequencing.
*KRAS expression was adjusted for 7 factors including age, T category, N category, M category, estrogen receptor, progesterone receptor, and HER2.
Correlation analyses among mRNA expression, CNA, and methylation of KRAS
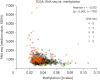
In TCGA dataset, the mRNA expression was higher when the value of CNA non increased (Figure 3A). The expression of mRNA was positively correlated with the value of CNA_lin (r = 0.577, p < 0.001, Figure 3B). In METABRIC dataset also, the mRNA expression was higher when the value of CNA non increased (Figure 3C). The expression of mRNA was positively correlated with CNA non (ρ = 0.343, p < 0.001, Figure 3D). In TCGA dataset, the mRNA expression was negatively correlated with methylation (r = 0.272, p < 0.001, Figure 4). Positive correlation was observed between mRNA expression and CNA_lin. Methylation showed negative correlation with both mRNA expression and CNA_lin (Supplementary Table 5).
Figure 3
Boxplots and scatter plots for correlation between mRNA expression and CNA of KRAS. Boxplots depicting the correlation between RNA seq and nonlinear data of CNA (A), and scatter plot showing the correlation between RNA seq and linear data of CNA (B) using TCGA database. Boxplots depicting the correlation between RNA mic and nonlinear data of CNA (C), and scatter plot showing the correlation between RNA mic and nonlinear data of CNA (D) using METABRIC database.
CNA = copy number alteration; RNA seq = RNA sequencing; TCGA = The Cancer Genome Atlas; RNA mic = RNA microarray; METABRIC = Molecular Taxonomy of Breast Cancer International Consortium; CNA_lin = copy number alteration linear; CNA non = copy number alteration nonlinear; GISTIC = Genomic Identification of Significant Targets in Cancer; RSEM = RNA sequencing by expectation maximization.
*p values by t-test between CNA = −2 and the other groups were 0.890 (CNA = −1), 0.133 (CNA = 0), 0.039 (CNA = 1), and < 0.001 (CNA = 2); †p values by t-test between CNA = −2 and the other groups were 0.513 (CNA = −1), 0.218 (CNA = 0), 0.044 (CNA = 1), and 0.069 (CNA = 2).

Analyses of KRAS mutation
In TCGA dataset, 6 (0.61%) out of 977 samples showed KRAS mutations. All the 6 were missense mutations. In METABRIC dataset, 12 (0.64%) out of 1,871 samples showed KRAS mutations, all of which were missense mutations. Details are described in Supplementary Table 6.
DISCUSSION
In 1964, Jennifer Harvey reported that a preparation of murine leukemia virus, later unveiled to carry Hras oncogene, caused sarcoma in newborn mice. It was the first report in the history of RAS research [1819]. In 1970, Werner Kirsten reported the existence of Kirsten murine sarcoma virus, later shown to carry Kras oncogene, by serial passage of murine leukemia viruses in Wister-Furth rats [20]. In 1982, human nucleotide sequences of HRAS and KRAS oncogenes from T24 and EJ bladder carcinoma cell lines were finally reported [2122]. Currently, three RAS members (HRAS, KRAS, and NRAS) are clinically the most notable founding members of the RAS subfamily. The RAS superfamily comprises over 150 members [23]. The most characterized RAS signaling pathway is the RAS-RAF-MEK-ERK cascade, which is also known as MAPK/ERK pathway. RAS can also activate several other effector pathways, including PI3K, RalGEF-Ral, and PLCε pathways [1]. Recent studies have reported that there are almost 140 genes whose intragenic mutations can contribute to human cancers [24]. More than 500 human cancer genes have been identified [6]. RAS genes are still the most frequently mutated oncogenes in human cancers followed by TP53, BRAF, and PIK3CA. KRAS mutation is dominant among all RAS subfamily members [6].
The results of this study revealed that breast cancer patients with high KRAS mRNA expression were significantly associated with worse survival regarding both OS and BCSS. The expression level of KRAS mRNA was a significant prognostic factor in breast cancer regarding OS according to both univariable and multivariable analyses. These findings were cross-validated by analyzing two different and independent datasets from TCGA and METABRIC databases. In METABRIC dataset, the group with high KRAS mRNA expression showed higher proportions of negative estrogen receptor, negative progesterone receptor, and positive HER2. As a result, the group with high KRAS mRNA expression showed lower proportion of luminal A subtype but higher proportions of other subtypes. The expression level of KRAS mRNA was a significant prognostic factor by univariable analysis. It remained an independent significant factor after adjusting for main clinicopathologic factors including estrogen receptor, progesterone receptor, HER2, and subtype. This is the first study to reveal the association between mRNA expression of KRAS and breast cancer prognosis. Few studies have reported the prognostic role of mRNA or protein expression of KRAS in human malignancies. A previous study has investigated the prognostic role of KRAS expression using RNA seq data of 1,017 patients with non-small cell lung cancer enrolled in TCGA program [25]. It reported that KRAS mRNA expression per se was not significantly correlated with OS. However, mRNA expression of surrogate signature genes for KRAS deletion was significantly associated with OS (high vs. low expression: HR, 2.3; 95% CI, 1.8–2.9) [25].
In this study, the prognostic impact of KRAS mRNA expression was effective only for luminal A subtype. Since there were no significant differences in clinicopathologic features between the two subgroups of luminal A according to mRNA expression level, the difference in KRAS mRNA expression level could be a crucial factor to differentiate the two. KRAS mRNA expression was significantly lower in luminal A compared to that in other subtypes. This could partly explain the favorable prognosis of luminal A subtype among all breast cancer subtypes. Although little is known about the prognostic role of KRAS mRNA expression in subtypes of breast cancer, some studies have reported the association of KRAS with endocrine resistance in luminal breast cancer subtype [9]. One study has shown that endocrine-resistant advanced breast tumors harbor oncogenic mutations in the MAPK signaling pathway including KRAS. These mutations are typically not detected in pre-treatment of primary tumors [8]. A genome-wide functional screening study has reported a gene set including KRAS whose silencing can cause sensitivity to endocrine therapy in breast cancer [11]. Another study has analyzed three independent gene expression datasets from GEO database and reported that RAS pathway activation is strongly associated with poor survival of patients having luminal breast cancers [26]. Further studies are needed to validate the prognostic role of KRAS in each breast cancer subtype. Some studies have reported the association between KRAS and breast cancer subtypes. One study has demonstrated that KRAS has a crucial role in the maintenance of mesenchymal phenotypes and metastatic ability of basal-type breast cancer by molecular experiments using breast cancer cell lines [27]. TCGA Research Network has reported that many components of the PI3K and RAS-RAF-MEK pathways are amplified, but not typically mutated, in basal-like breast cancer subtypes, including PIK3CA (49%), KRAS (32%), BRAF (30%), and EGFR (23%) [9].
In the present study, mRNA expression was positively correlated with CNA status but negatively correlated with methylation status. The positive correlation between mRNA expression and CNA was cross-validated using TCGA and METABRIC datasets while the negative correlation between mRNA expression and methylation was only proved in the TCGA dataset due to the lack of methylation data in METABRIC dataset. Little is known about the molecular regulation of KRAS at DNA, RNA, and protein levels, and others in breast cancer. TCGA Research Network has reported the landscape of genetic alterations in 10 canonical oncogenic signaling pathways including the RTK-RAS pathway using mutations, CNA, mRNA expression, gene fusions, and DNA methylation from 9,125 human malignancies profiled by TCGA. KRAS was the most frequently altered gene (9%), followed by BRAF (7%) and EGFR (4%) across all human cancers. KRAS alterations were the most commonly observed alterations in pancreatic carcinoma (72%), genomically stable colorectal cancer (69%), and lung adenocarcinoma (33%) [28].
Breast invasive carcinoma has been reported to have a rare frequency of KRAS mutation (0.7%), although it is the most common one in pancreatic ductal adenocarcinoma (97.7%) followed by colorectal adenocarcinoma (44.7%) and lung adenocarcinoma (30.9%) [6]. This study showed similar KRAS mutation frequencies (0.61% and 0.64% in TCGA and METABRIC datasets, respectively). Other studies have also reported rare frequencies of KRAS mutation in invasive breast cancers [529]. A previous study has shown that frequencies of KRAS mutations as: G12 (83%), G13 (14%), Q61 (2%), and others (1%) [630]. In the present study, G12 mutation accounted for 83.3% (5 out of 6 in a TCGA dataset and 10 out of 12 in a METABRIC dataset), similar to the previous report. Some previous studies have shown that rs61764370, an inherited variant residing in a KRAS 3' UTR microRNA binding site, is associated with increased risk of ovarian cancer and breast cancer and increased risk of reduced survival [1213]. Ovarian Cancer Association Consortium, Breast Cancer Association Consortium, and Consortium of Modifiers of BRCA1 and BRCA2 have revealed that rs61764370 is not associated with risk of ovarian or breast cancer or clinical outcome for patients with these cancers by analyzing data of 140,012 women enrolled in these consortia [14]. In the present study, KRAS mutation seemed to have little association with mRNA expression having a mutation frequency of approximately 0.6%.
This study has some limitations. First, we could not analyze the role of KRAS protein expression because of insufficient information in TCGA and METABRIC databases. The TCGA dataset has information on protein expression for only 74 subjects (6.8%) while the METABRIC dataset has no information regarding protein expression. Second, although this study revealed the prognostic role of the level of KRAS mRNA expression in breast cancer, the roles of other related genes were not analyzed. Third, we could not analyze the recurrence or metastasis pattern according to the level of KRAS mRNA expression due to lack of information. Further studies are needed to elucidate associations between KRAS mRNA expression and breast cancer prognosis.
In conclusion, high level of KRAS mRNA expression was associated with worse prognosis compared to low level of KRAS mRNA expression in breast cancer regarding both OS and BCSS. The expression level of KRAS mRNA was an independent significant prognostic factor in breast cancer regarding OS. These findings were cross-validated by analyzing two different and independent datasets from TCGA and METABRIC databases. The prognostic role of KRAS mRNA expression was effective only for luminal A subtype, which showed significantly lower level of mRNA expression compared to that in other subtypes. The expression level of mRNA was positively correlated with CNA status but negatively correlated with methylation status. The expression of mRNA had little association with mutation status in breast cancer having a mutation frequency of approximately 0.6%. Further studies are needed to validate the prognostic role of KRAS mRNA expression in breast cancer, thus paving a way for clinical application of KRAS in practice.
 XML Download
XML Download