

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Breast cancer, which ranks first among female malignant tumors, has become a major public health problem [1]. The prognosis of patients with breast cancer depends on the presence of metastasis and recurrence, which largely depend on the presence of breast cancer stem cells [23]. Identifying tumor stem cells in tumor tissue is important for the diagnosis and treatment of tumors. Breast cancer stem cell-specific surface molecules and their abnormal activation signaling pathways distinguish them from other tumor cells and also serve as the identification points for the separation of cancer stem cells [4].
Studies on molecular markers of breast cancer have been widely conducted and the findings of these studies are being applied in clinical diagnosis and treatment. For example, aldehyde dehydrogenase 1 family member A1 (ALDH1) has been considered a poor prognostic factor in breast cancer [5]. The multifunctional receptor cluster of differentiation (CD) 44, which mediates cell–cell and cell–matrix interactions, is highly expressed and functions in breast cancer stem cells. Conversely, the expression of membrane protein CD24, which is highly expressed in malignant tumors, is lower in breast cancer stem cells. CD44+/CD24- expression characteristics can be used in combination for isolation of breast cancer stem cells [6]. In addition, some embryonic stem cell marker molecules, such as organic cation/carnitine transporter4 (Oct4), Nanog, and Bmi-1, are also frequently expressed in breast cancer stem cells. Cancer stem cell marker Bmi-1 expression is associated with basal-like phenotype and poor survival in breast cancer [7]. Biotargeted clearance of cancer stem cells is suggested to be the best method to completely cure malignant tumors. For example, RG7356, a monoclonal antibody drug that specifically targets CD44 molecules, can be expected to kill breast cancer stem cells due to its specific effect on CD44-positive tumors [8]. However, currently, there are few available drugs that target breast cancer stem cells. The recognition of reliable markers to distinguish stem cells in breast cancer will be decisive for the development of specific targeted therapies.
Several important intracellular signaling pathways, such as the Notch, Hedgehog, and Wnt signaling pathways, are common terminal pathways through which molecules regulate cancer stem cells [910]. Activating intracellular Notch signaling could upregulate the expression of specific surface markers of cancer stem cells, whereas blocking this signal can cause apoptosis in cancer stem cells [11]. Hedgehog signaling inhibitors can reverse the resistance of cancer stem cells to chemotherapy drugs [12]. The Wnt signaling pathway plays an important role not only in normal embryonic cells, but also in malignant stem cells, such as regulating proliferation, inducing morphological changes, and determining cell differentiation [13]. The extensive association of the Wnt pathway with the Notch and Hedgehog pathways further confirms the special importance of the Wnt pathway in the development of tumorigenesis [14]. In recent years, it has been found that the Wnt/β-catenin signaling pathway is abnormally activated in cancer stem cells. In gastric cancer spheres, the expression of β-catenin is significantly increased [15]. In breast cancer, targeting Wnt signaling not only inhibits the growth of breast cancer stem cells but also helps reduce the aggressiveness of triple-negative breast cancer via microRNA regulation in vitro and in vivo [16]. These studies indicate that the Wnt/β-catenin signaling is important for self-renewal of breast cancer stem cells.
Our present study demonstrated that endogenous KIF11 promotes the self-renewal capacity of breast cancer cells and enhances the characteristics of breast cancer stem cells by participating in the activation of the Wnt/β-catenin signaling pathway. We determined that KIF11 plays an important role in the tumorigenicity and recurrence of human breast cancer and can be expected to serve as an effective therapeutic target in breast cancer.
METHODS
Ethics statement
Female nonobese diabetic/severe combined immunodeficiency disease (NOD/SCID) mice (4–5 weeks old, 18–20 g) were purchased from Hunan SJA Laboratory Animal Co. Ltd (Changsha, China). The Institutional Animal Care and Use Committee of Guang Dong Pharmaceutical University (gdpulac2018049) approved all experimental procedures. All experiments with animals were conducted in accordance with the guidelines described in the National Institutes of Health “Guide for the Care and Use of Laboratory Animals”.
Microarray data processing and visualization
The RNA sequencing datasets downloaded on November 29, 2014 from The Cancer Genome Atlas (TCGA) contain samples of 1,092 breast cancer tissues and 111 adjacent normal breast tissues. In 1,092 cases of breast cancer, there were 63 cases of luminal A subtype, 398 cases of luminal B subtype, 165 cases of human epidermal growth factor receptor 2 (HER2) overexpression subtype, 122 cases of basal subtype, and 339 cases of normal subtype. Profile data were extracted using Excel and MEV 4.9 (http://www.tm4.org/mev). Gene concentration analysis was performed using gene set enrichment analysis (GSEA 2.2.1; http://www.broadestitute.org/gsea).
Cell lines, plasmids, and real-time polymerase chain reaction (RT-PCR)
Breast cancer cell lines were cultured and established as previously described [17]. For depletion of KIF11, 2 human small interfering RNA sequences were cloned into pSuper-retro-puro, as previously described [18]. Throughout the entire procedure, total RNA extraction, reverse transcription, and RT-PCR were conducted as previously described [17]. All target genes were homogenized with the housekeeping gene glyceraldehyde 3-phosphate dehydrogenase (GAPDH) to control the variability of expression levels and were calculated as 2−[(CT of indicated genes)–(CT of GAPDH)], where CT represents the threshold cycle for each transcript. The primers used were as follows: KIF11-forward, 5′-TATTGAATGGGCGCTAGCTT-3′; KIF11-reverse, 5′-TCGTCTGCGAAGAAGAAAGA-3′; c-Myc-forward, 5′-TTCGGGTAGTGGAAAACCAG-3′; c-Myc-reverse, 5′-CAGCAGCTCGAATTTCTTCC-3′; cyclin D1 (CCND1)-forward, 5′-AACTACCTGGACCGCTTCCT-3′; CCND1-reverse: 5′-CCACTTGAGCTTGTTCACCA-3′; Oct4-forward, 5′-GGTTCTCGATACTGGTTCGC-3′; Oct4-reverse, 5′-GTGGAGGAAGCTGACAACAA-3′; ALDH1-forward, 5′-AGGGGCAGCCATTTCTTCTCA-3′; ALDH1-reverse, 5′-CACGGGCCTCCTCCACATT-3′; CD44-forward, 5′-ACCCCATCCCAGACGAAGACAGTC-3′; CD44-reverse, 5′-GGGATGAAGGTCCTGCTTTCCTTCG-3′; Nanog-forward: 5′-ATGGAGGAGGGAAGAGGAGA-3′; Nanog-reverse, 5′-GATTTGTGGGCCTGAAGAAA-3′; GAPDH-forward, 5′-GACTCATGACCACAGTCCATGC-3′; GAPDH-reverse, 5′-AGAGGCAGGGATGATGTTCTG-3′.
Western blotting
Western blot analysis was performed according to standard methods [17], using antibodies against KIF11 (Santa Cruz Biotechnology, Inc., Dallas, USA), and β-actin (Sigma, Saint Louis, USA) was detected as a loading control.
Mammosphere culture
As mentioned by Song et al. [17], 1,000 single cells were seeded in 24-well culture plates with low adsorption. Suspension culture was conducted using 500 μL serum-free Dulbecco's Modified Eagle Medium: Nutrient Mixture F-12 (DMEM-F12, Sigma). A total 50 μL of culture medium was added every day. On day 20, qualitative and quantitative values of mammospheres were obtained using Zeiss AxioVision software (Carl Zeiss Co. Ltd., Jena, Germany).
Hoechst 33342 staining and flow cytometry
To identify and isolate side-population (SP) cells, the cells were dissociated and resuspended at 1,000,000 cells/mL in DMEM supplemented with 5% fetal bovine serum, preincubated at 37°C for 30 minutes with or without 100 mM verapamil (Sigma). Next, 2.5 mg/ml Hoechst 33342 (Sigma) was added to this media with 20 minutes of continuous processing, and then incubated at 37°C for 90 minutes. Following this, these cell mixtures were incubated on ice for 10 minutes and washed twice with ice-cold phosphate buffered saline. The mixtures were then analyzed (20,000 cells per experiment with 3 replication) using a FACStar plus (Becton Dickinson Immunocytometry Systems, San Jose, USA) cell sorter equipped with dual Coherent I-90 lasers.
Xenograft tumor model
Based on previous explorations, 30 mice were divided into 3 groups and transplanted with 10,000 cells, 5,000 cells and 1,000 cells, respectively. The mammary fat pads of each mouse were injected with SKBR-3-Ri-Vec cells on the left side and with SKBR-3-RNAi1 cells on the right side. On day 28, the animals were euthanized and the tumors were excised, photographed, and counted.
Immunofluorescent assay
Cell fixing and staining were conducted, as described previously [19]. The final working dilution of anti-human β-catenin monoclonal antibody (Abcam, Cambridge, UK) used in cell staining was 1:200.
We used 4′,6-diamidino-2-phenylindole (DAPI; Sigma Chemical, St. Louis, USA) staining to identify the nuclei. Finally, we used laser scanning microscopy (Axioskop 2 Plus; Carl Zeiss Co. Ltd.) to obtain and analyze the images.
Luciferase reporter assay
Cells were inoculated into 24-well plates for normal culture. Twenty-four hours later, the identified plasmids plus 1 ng of the pRL-TK Renilla plasmid were transfected into the cells using Lipofectamine 2000 reagent (Life Technologies, Carlsbad, USA). After 2 days, the Dual-Luciferase Reporter Assay (Promega, Madison, USA) was used for analysis, according to the manufacturer's instructions. All experiments were performed in triplicate.
RESULTS
Upregulated KIF11 positively affects stem cell-regulating genes
Our previous studies have shown that KIF11, which is upregulated in breast cancer, is closely related to the clinicopathological features of breast cancer [18]. Herein, we validated our previous findings by analyzing data from TCGA. After being grouped and compared, KIF11 was upregulated in 1,092 malignant breast tissues (Tumor) compared with that in 111 paracancerous normal breast tissue samples (Normal; Figure 1A). As shown in Figure 1B, KIF11 was upregulated in all subtypes of breast cancer, as compared with normal breast tissue (p < 0.05). Except for the comparison between HER2 overexpression and basal-like subtypes, there was a statistically significant difference in the expression of KIF11 between any 2 subtypes. Moreover, the KIF11 expression from TCGA profiles was categorized and analyzed using GSEA to investigate the significance of KIF11 upregulation in breast cancer. According to the detection value of KIF11 expression, 1,092 patients were divided into a high-expression group (KIF11-H) and low-expression group (KIF11-L; the overall average was the cutoff value). As shown in Figure 1C, the KIF11 level was positively associated with the stem cell-positive regulatory gene, indicating its potential to promote breast cancer self-renewal.
Figure 1
Upregulated KIF11 positively affects stem cell-regulating genes. (A) The RNA level of KIF11 in 1,092 breast cancer tissues (Tumor) compared with 111 adjacent normal bladder epithelial samples (Normal) in the TCGA profile validated the upregulation of KIF11 in breast cancer. (B) The RNA level of KIF11 was upregulated in every subtype of breast cancer. (C) Gene set enrichment analysis plots of TCGA data show KIF11 levels positively affected stem cell-regulating genes.
KIF11 = kinesin family member 11; SEM = standard error of the mean; TCGA = The Cancer Genome Atlas; HER2 = human epidermal growth factor receptor 2; NS = nonsense.
*p < 0.05; †p < 0.001.

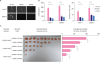
Silencing KIF11 reduces the proportion of stem cells in breast cancer cell lines
To investigate the biological role of KIF11 in breast cancer stem cell self-renewal, endogenous KIF11 in MCF-7 and SKBR-3 cells was stably silenced. As shown in Figure 2A and B, messenger RNA (mRNA) levels and protein expression of KIF11 were significantly reduced in KIF11-silenced cells. As shown in Figure 2C, the mRNA expression of cancer stem cell markers (Oct4, Nanog, ALDH1, and CD44) was significantly downregulated in KIF11-silenced cells. The proportion of SP cells (Hoechst 33342-effluxed cells, red) was significantly reduced in KIF11-silenced cells (Figure 2D). Verapamil-treated cells were used as negative controls. These results indicate a positive correlation between KIF11 and the proportion of stem cells in breast cancer.
Figure 2
Silencing KIF11 reduces the proportion of stem cells in breast cancer cell lines. (A) The mRNA expression level of KIF11 was successfully decreased in the indicated cells. (B) The protein expression of KIF11 was successfully decreased in the indicated cells. (C) The mRNA level of the indicated genes was detected using real-time polymerase chain reaction. (D) Flow cytometry analysis of Hoechst 33342-effluxed cells show that the proportion of SP cells was significantly decreased in the KIF11-silenced cells.
KIF11 = kinesin family member 11; mRNA = messenger RNA; Oct4 = organic cation/carnitine transporter4; ALDH1 = aldehyde dehydrogenase 1; CD44 = cluster of differentiation 44.
*p < 0.05.
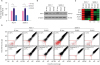
Silencing KIF11 attenuates the self-renewal ability of breast cancer cells
As shown in Figure 3A, silencing KIF11 reduced mammosphere formation in MCF-7 and SKBR-3 cells. Summarized in Figure 3B, the expression of KIF11 was positively correlated with the size and number of mammospheres. Furthermore, when subcutaneously implanted into NOD/SCID mice, KIF11-silenced SKBR-3 cells formed fewer and smaller tumors than vector-transfected SKBR-3 cells (Figure 3C), indicating that the proportion of stem cells in the KIF11-silenced SKBR-3 cells was lower than that in the vector-transfected cells. Collectively, these assays demonstrated that KIF11 is positively correlated with the self-renewal ability of breast cancer cells.
Figure 3
Silencing KIF11 attenuates the self-renewal ability of breast cancer cells. (A, B) Representative micrographs and colony numbers in the mammosphere culture assay show that silencing KIF11 reduced mammosphere formation in MCF-7 and SKBR-3 cells. (C) Images of excised tumors from nonobese diabetic/severe combined immunodeficiency disease mice at 28 days after injection with the indicated cells show that KIF11-silenced SKBR-3 cells formed fewer and smaller tumors than vector-transfected SKBR-3 cells.
Each bar represents the mean±standard error of the mean of 3 independent experiments.
KIF11 = kinesin family member 11.
*p < 0.05; †p < 0.001.
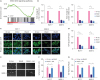
Silencing KIF11 inactivates Wnt/β-catenin signaling
Using in-depth GSEA, we preliminarily identified the intracellular signaling pathway of biological processes mediated by KIF11. As shown in Figure 4A, the expression of KIF11 was positively correlated with the gene characteristics of Wnt activation, suggesting that Wnt signaling may be the pathway associated with KIF11. RT-PCR showed that transcription factor T-cell factor 1 (TCF1) and CCND1, the key downstream genes of the Wnt signaling pathway, decreased sharply in KIF11-silenced cells (Figure 4B). In both MCF-7 and SKBR-3 cells, a large proportion of β-catenin is distributed in the nucleus, which also suggests that β-catenin is activated and is fulfilling its role in promoting downstream gene transcription. However, β-catenin was almost evenly distributed in the cytoplasm and membrane of KIF11-silenced cells, indicating that the Wnt/β-catenin signaling pathway was in a relatively silent state (Figure 4C). Analysis of the β-catenin reporter gene, based on TCF4 and LEF1 genes, showed that silencing KIF11 significantly reduced the activation activity of β-catenin in cells (Figure 4D). Moreover, Wnt agonists significantly neutralized the inhibition of KIF11 downregulation on mammosphere formation and intracellular β-catenin-mediated transcriptional activity (Figure 4E-G). Altogether, these data suggest that KIF11 enhances the nuclear translocation of β-catenin, thus activating the classical Wnt signaling pathway.
Figure 4
Silencing KIF11 inactivates Wnt/β-catenin signaling. (A) Gene set enrichment analysis plots of The Cancer Genome Atlas data show that the expression of KIF11 was positively correlated with the gene characteristics of Wnt activation. (B) Real-time polymerase chain reaction analysis shows that TCF1 and CCND1 expression decreased sharply in KIF11-silenced cells. (C) Immunofluorescence staining show distribution of β-catenin from nucleus to cytoplasm in KIF11-silenced cells (×400). (D) Analysis of the β-catenin reporter gene, based on TCF4 and LEF1 genes, showed that silencing KIF11 significantly reduced the activation activity of β-catenin in cells. (E, F) Representative micrographs and colony numbers in the mammosphere culture assay show that Wnt agonists neutralized the inhibition of KIF11 downregulation in mammosphere formation. (G) Luciferase reporter assay of TCF/LEF transcriptional activity shows that Wnt agonists significantly neutralized the inhibition of KIF11 downregulation on intracellular β-catenin-mediated transcriptional activity.
KIF11 = kinesin family member 11; DAPI = 4′,6-diamidino-2-phenylindole; TCF = transcription factor T-cell factor; LEF = lymphoid enhancer-binding factor; CCND1 = cyclin D1; mRNA = messenger RNA.
*p < 0.05.
DISCUSSION
KIF11 is a mitotic kinesin that plays a crucial role in the formation of bipolar mitotic spindles by hydrolyzing ATP to push apart anti-parallel microtubules [20]. KIF11 has been reported to be involved in the progression of many cancers; for example, in renal clear cell carcinoma tissues, the level of KIF11 mRNA is higher than that of corresponding noncancerous tissues; in esophageal cancer and colorectal cancer cells, the expression of KIF11 is downregulated, significantly reducing stem cell-associated spheroid formation. In oral cancer cells, the use of KIF11 inhibitors significantly inhibits cell proliferation and induces apoptosis [212223]. Our previous study demonstrated that the expression of KIF11 was markedly overexpressed in human breast cancer cells and breast cancer tissues. According to the expression and prognosis of estrogen receptor, progesterone receptor, and HER2, breast cancer can be divided into the following 5 subtypes: luminal A, luminal B, HER2 overexpression, basal, and normal [24]. These molecular subtypes, which more accurately reflect the biological characteristics of breast cancer, can be used to assess prognosis and to select more targeted personalized treatments [25]. In our present study, KIF11 was found to be upregulated in all subtypes of breast cancer, as compared with normal breast tissue. Except for the comparison between HER2 overexpression and basal-like subtypes, there was a statistically significant difference in expression of KIF11 between any 2 subtypes. This result indicates that the expression level of KIF11 can be used as a joint factor in molecular typing of patients, to help develop individualized and systematic treatment options.
We previously reported that statistical analysis showed a significant correlation between upregulation of KIF11 expression and distant metastasis/prognosis in patients with breast cancer [18]. Multivariate analysis revealed that KIF11 upregulation may be an independent prognostic indicator for the survival of patients with breast cancer. Because breast cancer recurrence and metastasis are closely related to the existence of breast cancer stem cells, we speculated that KIF11 may be involved in the regulation of breast cancer stem cells. In our present study, we validated our previous findings through an analysis of data collected in the TCGA profile and found a positive correlation between KIF11 levels and stem cell-regulating genes. Flow cytometry showed that the proportion of SP cells was significantly reduced in KIF11-silenced cells. Mammosphere formation assays and tumor implantation experiments in NOD/SCID mice both demonstrated that the expression of KIF11 was positively correlated with self-renewal of breast cancer cells. These studies suggest that endogenous KIF11 could increase the proportion of stem cells in breast cancer cell lines and enhance the self-renewal ability of breast cancer cells.
Wnt signaling pathways are classified into 4 categories, based on their different functions [26]. The canonical Wnt/β-catenin signaling pathway is the most relevant to tumor regulation [27]. As a key molecule in the canonical Wnt/β-catenin signaling pathway, β-catenin protein is often phosphorylated and degraded by binding to proteins such as Axin and GSK3β in the cytoplasm; unphosphorylated β-catenin proteins are often localized in the nucleus and activate the downstream signal [28]. Inhibition of the Wnt/β-catenin signaling pathway may be effective in inhibiting the metastasis and recurrence of breast cancer [16]. Herein, we found that silencing KIF11 inactivates the Wnt/β-catenin signaling pathway by changing the intracellular localization of β-catenin. Moreover, Wnt agonists significantly neutralized the inhibition of KIF11 downregulation in mammosphere formation and intracellular β-catenin-mediated transcriptional activity. The regulation of this key signaling pathway also illustrates the important role of KIF11 in breast cancer stem cells and its potential value as a therapeutic target.
In summary, the results of the present study indicate that endogenous KIF11 regulates the stem cell-associated progress by activating the Wnt/β-catenin signaling pathway, suggesting a potential therapeutic target role of KIF11 in breast cancer.
 XML Download
XML Download