

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Endogenous new blood vessel formation and microvascular angiogenesis are important self-defense mechanisms when tissue is exposed to severe ischemia.1) However, these mechanisms; endogenous collateral vessel formation and capillary angiogenesis; are mainly mediated by endothelial cell (EC) proliferation and migration, and thus, are often hampered by co-existing morbidity such as diabetes mellitus, smoking, hypercholesterolemia, etc.2) It is not rare for diabetic patients to lose some parts of their extremities after suffering from severe peripheral artery obstructive disease (PAD) because of insufficient development of collateral vessels and angiogenesis. Therefore, the treatment option called “therapeutic angiogenesis” is an important strategy to salvage tissues against critical ischemic conditions.1)
Subcutaneous adipose tissue can be harvested by relatively easy and less invasive methods, an established liposuction method. Furthermore, recent studies have indicated that subcutaneous adipose tissues contain stem/progenitor cells that can give rise to several lineage cells including fat, bone, cartilage, muscle, and etc.3)4) These progenitor cells are termed as adipose-derived stem cells (ASCs or ADSCs), or adipose-derived regenerative cells (ADRCs), and these terminologies including adipose-derived stromal vascular fraction (SVF) are essentially considered as an identical cell fraction.5)6) ADSCs/ADRCs can also release multiple angiogenesis-promoting growth factors such as vascular endothelial growth factor (VEGF), hepatocyte growth factor (HGF), fibroblast growth factor (FGF), and chemokine stromal cell-derived factor-1 (SDF-1).7)8)9) The combination of these biological properties (i.e., mesenchymal progenitor cell supply and angiocrine factors release) suggests that autologous subcutaneous adipose tissue will be a good candidate for a cell source of therapeutic angiogenesis.7)8)9)10)11)12)13)14)
THERAPEUTIC ANGIOGENESIS: GROWTH FACTORS AND CYTOKINE THERAPY
Past great professors, Drs. Judah Folkman and Jeffrey Isner, were pioneers for angiogenesis research. Isner and co-workers15)16)17) had initiated the concept of “therapeutic angiogenesis” in the 1980s. Folkman18)19) observed that angiogenesis was essential for the growth and metastasis of solid tumors. His original idea that the suppression of tumor angiogenesis would be effective against tumor growth had been developed into a new paradigm of anti-angiogenic therapy against neoplastic disorders, called “tumor-dormancy therapy”.18)19)
After the identification of angiogenic growth factors such as VEGF and basic FGF (bFGF), investigators in the cardiovascular field started testing their hypothesis that facilitating angiogenesis with growth factor genes and/or proteins would improve tissue blood flow and function in critically ischemic diseases.15)16)17)20) A number of preclinical animal studies together with in vitro laboratory data supported the safety and feasibility of clinical application of therapeutic angiogenesis using growth factor genes or proteins. Thereafter, abundant clinical trials have been performed to conduct therapeutic angiogenesis with genes and cytokines.21) Initial small clinical trials showed successful results, however, clinical studies with randomized placebo-controlled trails failed to show sufficient improvement of angiogenesis, tissue function and patients' symptoms.21) These findings suggested that the clinical trial of therapeutic angiogenesis using single factor might have a limited efficacy. This will be explained by the fact that the expression of more than 750 genes were either up-regulated or down-regulated more than 2-fold by acute ischemic event in vivo,22) and such dramatic changes may not easily be overcome by a single cytokine treatment. So far, only small numbers of studies using HGF or bFGF have been shown to reveal promising data, but these may be because a number of cytokines or transcription factors are expressed by downstream to these cytokine-mediated signal transduction.23)24)25)26)
THERAPEUTIC ANGIOGENESIS: CELL THERAPY
A pioneering work by Drs. Asahara et al.27) regarding the identification of endothelial progenitor cells (EPCs) in human peripheral blood (PB) was published in 1997. Since EPCs and hematopoietic stem cells (HSCs) share several cell surface antigens such as Flk-1, Tie-2, CD31, and CD34, EPCs are believed to derive from hemangioblasts or closely related cells existing in the bone marrow (BM) in adults.28) Circulating EPCs derived from the BM participate in new blood vessel formation and/or re-endothelialization on the injured arterial wall.27) Although transplantation of culture expanded EPCs could successfully augment angiogenesis and tissue blood supply in experimental animal models,29) this procedure has not been developed into clinical trials because of difficulties in culture expansion techniques and limited number of cells to implant.
Instead, based on the fact that patients' own BM can be obtained by an established way and that the fresh BM contains HSCs and EPCs, implantation of autologous BM mononuclear cells has been investigated in both preclinical and clinical studies.30)31)32)33) Adult BM has been shown to contain EPCs that mobilize into peripheral circulation after stimulation by ischemia or by cytokines such as VEGF and SDF-1.34)35)36) We previously demonstrated that implantation of autologous BM mononuclear cells into ischemic skeletal muscles successfully augmented angiogenesis and collateral vessel formation in both animal studies and human trials.30)31)33)37)38)39)40) Especially in the clinical study called the “Therapeutic angiogenesis with cell transplantation (TACT) trial,” we have shown that more than 80% of Burger's disease patients and about 50% of PAD patients were well responsive to this therapeutic procedure.38)39)40)
However, there were several limitations to this procedure. First, the procedure itself is too invasive for patients having critical limb ischemia (CLI). In the TACT protocol, approximately 1–3×109 BM mononuclear cells were required for cell transplantation in 1 limb, which contains 1–3×107 CD34+ cells and needs about 800 mL of fresh BM aspirated from the ileum.33)37) Secondly, although the safety and efficiency of the TACT protocol have been established, we recently reported that patients with severe end-stage PAD had poor responses to the TACT procedure.37)39) For example, patients with diabetes or with chronic hemodialysis due to end stage renal disease (ESRD) showed limited responses to the TACT procedure.33) Moreover, several studies indicated that patients with severe PAD, ischemic heart disease and/or multiple coronary risk factors had diminished functions of EPCs and poor responses to angiogenic cell therapy.41)42)43)44) These results suggest that even autologous BM mononuclear cells were isolated, the functions of stem/progenitor cells are already reduced to induce sufficient level of angiogenesis after implantation into the “host” ischemic tissues. It has been demonstrated that the efficacy of the TACT procedure was limited in patients with poorly controlled diabetes and ESRD.33)39) In such patients, the number and functions of EPCs are also reduced and are difficult to be isolated. Also, we previously demonstrated that the number and function of circulating EPCs were markedly reduced in chronic smokers even without other risk factors.44)
ADIPOSE-DERIVED MESENCHYMAL STEM/PROGENITOR CELLS
Since the BM aspiration is invasive for patients with severe ischemic diseases, less invasive techniques for isolating other candidate cells for angiogenic therapy have been explored. One of the attractive sources for isolating cells for therapeutic angiogenesis would be autologous subcutaneous adipose tissue.3)4)5)6)7)8)9)10)11)12)13)14)45)46)47)48)49)50) Adipose tissue mainly comprises 2 classes of cells. One is mature adipocytes (MAs) forming major volume of the adipose tissue, and the other is stromal cells termed as SVF. Studies have demonstrated that SVF contains multipotent mesenchymal stem/progenitor cells that could differentiate into various lineage cells including adipocytes, fibroblasts, myocytes, pericytes, osteoblasts, chondrocytes, and etc.3)4) Recently, adipose tissue has been shown to contain multipotent mesenchymal stem cells termed adipose-derived stem/progenitor cells (ASCs or ADSCs) or ADRCs that have an ability to regenerate various damaged tissues. ADRCs could be isolated from even a small amount of human subcutaneous adipose tissues and culture-expandable.4) Autologous ADRCs could be isolated by minimal invasive technique such as a local liposuction method even in patients with CLI.
THERAPEUTIC ANGIOGENESIS USING ADIPOSE-DERIVED REGENERATIVE CELLS: A NEW PARADIGM
Previous studies showed that ADRCs could differentiate into ECs or EPCs,10)11) but in our investigation we could not confirm ADRCs to differentiate into the endothelial lineage cells.9) One study used low-serum conditioned medium supplemented with VEGF and insulin-like growth factor-1 to make ADRCs differentiation to ECs or EPCs.10) The difference of such culture conditions might have affected the differentiation capacity of ADRCs into ECs.47) Nevertheless, a recent study failed to show that human ADRCs differentiated into ECs even under culture with endothelial specific medium EC growth medium-2.48) In addition in vivo study, we could not confirm differentiation of ADRCs into ECs. Implanted GFP-positive ADRCs expressed no endothelial makers but CD140b, also know as PDGF receptor B and a myofibroblast-specific marker, and co-localized with vascular pericytes.9) These results suggest that ADRCs might not have capability to differentiate into EPCs or ECs but into pericytes or vascular smooth muscle cells after in vivo implantation. Several recent studies also demonstrated that ADRCs could differentiate into pericytes in vitro and in vivo.48)49)50)
Although our study and previous studies failed to show the ADRCs or ASCs gave rise to the ECs, it has been shown that ADRCs can secrete multiple angiogenic growth factors.7)8) Therefore, autologous ADRCs may be very useful for therapeutic angiogenesis via a growth factor-dependent paracrine mechanisms. Nakagami et al.8) and Sumi et al.13) previously reported that implanation of adipose-derived mesenchymal cells induced angiogenesis via a secretion of growth factors. We also found that implantation of ADRCs significantly augmented angiogenesis in a mouse model of severe hind limb ischemia (Figure 1).9) Our data indicated that implantation of ADRCs induced angiogenesis not only by angiogenic cytokine release but also by chemokines such as SDF-1.9) We found that implantation of MAs into skeletal muscles worsened angiogenesis compared to saline-injected control mice. We found that ADRCs expressed SDF-1, and the abundance of messenger RNA (mRNA) and protein expression was significantly greater in ADRCs than in MAs.9) Recent studies indicated that MAs release other adipocytokines including tumor necrosis factor-α and interleukin-6. These inflammatory cytokines might have negatively affected angiogenesis by MA implantation that was observed in our study.7) SDF-1 is a member of CXC chemokines originally isolated from murine BM stromal cells.51) CXCR4 is the receptor for SDF-1 and is a co-receptor for HIV type 1 infection.52) The SDF-1/CXCR4 axis regulates multiple physiological processes including embryonic development and organ homeostasis. SDF-1 is considered as one of the key regulators of EPCs mobilization from the BM into PB.36) Thus, SDF-1 has been shown to augment neovascularization by acceleration of EPCs recruitment into ischemic foci.36)53) In addition, VEGF is one of powerful angiogenic cytokines that can also mobilize EPCs from BM and inhibit EPC apoptosis.54) Interestingly, in the mouse ischemic hind limb model, VEGF-A-mediated angiogenesis partly depends on the activation of the SDF-1-CXCR4 pathway.53) Taken together, chemokine SDF-1 likely plays a pivotal role for the ADRC-mediated angiogenesis.9) In fact, therapeutic efficacies and mobilization of EPCs after ADRCs implantation was markedly suppressed by injection of an anti-SDF-1 neutralizing monoclonal antibody in our study.9)
Figure 1
Efficacy of autologous BM-MNCs implantation in patients with CLI. Kaplan-Meier analysis of overall survival, major amputation-free, and total amputation-free survival following BM-MNC implantation in patients with PAD (ASO), TAO, and CDV.40)
ASO = arteriosclerosis obliterans; BM-MNC = bone marrow mononuclear cell; CDV = collagen disease-associated vasculitis; CLI = critical limb ischemia; PAD = peripheral artery disease; TAO = thromboangiitis obliterans.

ADIPOSE-DERIVED REGENERATIVE CELL THERAPY FOR ANGIOGENESIS: CLINICAL TRIALS
Previously, Lee et al.55) conducted a clinical trial to test the safety and efficacy of implanting autologous ADRCs into the ischemic limbs of patients with Burger's disease and diabetes. Their study involved 15 patients with CLI (12 with Burger's disease and 3 with diabetic foots). ADRCs were isolated from subcutaneous adipose tissues, expanded in culture until passage 3, and intramuscularly injected into the ischemic muscles of patients. They showed that this cell-based therapy using ADRCs might be feasible and effective for increasing blood flow and improving pain and clinical outcomes in patients with CLI.55)
Although the efficacy of autologous ADRC implantation for therapeutic angiogenesis has been established in animal and human studies, it is still unknown how many cells should be used for optimal angiogenesis. In the case of BM mononuclear cells, it has been reported that the injection of too many cells resulted in adverse effects in an animal model.56) The optimum dosage of ADRC implantation will be necessary to be determined. Second question is what the optimal cell isolation method is. Recently, freshly isolated ADRCs by means of collagenase digestion followed by an automatic centrifuge system using a specific machine is useful for regeneration of adipose tissues for reconstructive surgeries.46)57) Such technique needs neither cell cultivation nor an artificial cell processing. If 2 classes of ADRCs obtained by either culture method or fresh isolation method have equal potency in terms of releasing angiogenic growth factors, it will be useful to employ such machine for cell-mediated therapeutic angiogenesis. Thirdly, either ADRC number or functions may be affected by concomitant patients' conditions or atherogenic risk factors. For example, SDF-1 expression is considered to be reduced in diabetes,58) and the function and number of progenitor cells are generally reduced in patients with multiple risk factors including diabetes, hypertension, aging and smoking.41)42)43)44)59)60) Such pathological conditions are commonly complicated in patients with PAD. Little is known whether ADRC number and functions are influenced by the presence of such risk factors as well. Before applying this new technique to therapeutic angiogenesis, it will be necessary to elucidate these points.
POTENTIAL NEW MECHANISMS OF ADIPOSE-DERIVED REGENERATIVE CELL-MEDIATED ANGIOGENIC THERAPIES
Although direct differentiation of cells and cytokine/chemokine released from ADRCs have been considered as major mechanisms for therapeutic angiogenesis,8)9)61)62)63)64) a recent progress in science has identified additional potential mechanisms for ADRC-mediated angiogenic therapy.65) One of these mechanisms may be extracellular microvesicles (EMVs) released from ADRCs.66) Recently, a variety of cells have been shown to release EMVs that contains proteins, lipids, mRNAs, precursor microRNAs (miRNAs), miRNAs, transfer RNAs, and etc (Figure 2). Among them, ADRCs have been shown to release miRNAs to stimulate angiogenesis or tissue regeneration. Kang et al.66) showed that ASCs or ADSCs induce angiogenesis by EMV transport of miRNA-31. We recently reported that cardiac implanted ADRCs released miRNA-214 that was taken up by cardiomyocytes by a clathrin-mediated endocytosis.67) These EMV-transported miRNA-214 protected cardiomyocytes against acute myocardial infarction in a mouse model. These content molecules of EMVs released from mesenchymal stem/progenitor cells gained more and more attention for future application of cell-free conditioned medium-mediated regenerative medicine.65)
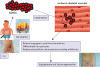
Figure 2
Mechanisms for the ADRCs-mediated angiogenesis in ischemic tissues.
Subcutaneous adipose tissue is isolated from patient with CLI. ADRCs are isolated by the adipose tissues and directly implanted into the patient's own ischemic skeletal muscles. Implanted ADRCs release angiogenic cytoines/chemokines, extracellular microvesicles including miRNAs that stimulate local angiogenic response. A part of ADRCs differentiated into pericytes and support the angiogenesis.
ADRC = adipose-derived regenerative cell; CLI = critical limb ischemia; miRNA = microRNA.
 XML Download
XML Download