

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
The approximately 0.2%~2% of the world population suffer from androgenetic alpopecia12. Hair loss occurs due to psychological and physical stress, and dandruff34. So far, minoxidil and finasteride was approved as a drug against for the hair loss567. Minoxidil is an anti-hypertensive, has been shown to stimulate hair growth in the treatment of vasodilatation on body8; finasteride was promoted hair growth in androgenetic alopecia (AGA) with male patients9. However, these drugs have unpredicatable efficacy and side effects, leading to limited therapeutic use10. Therefore, effective AGA treatment agents should be further developed. Recently, many researchers have reported natural extracts that promote hair growth1112.
Melandrium firmum (MF) is a herbal plant used to treat gonorrhe, anuria and breast cancer and widely distributed in Korea1314. However, its effect on hair growth has not been reported. The aim of this study was to investigate effects of MF on the prevention or treatment of AGA by selectively inhibiting 5α-reductase activity and its action mechanisms. Our study demonstrated that topical MF extract promotes hair growth in C57BL/6J mice.
MATERIALS AND METHODS
Preparation of MF
Whole plants of MF were purchased from Deakwang, Chuncheon, and the plant identification was confirmed by Emeritus Professor Heung Jun Chi (Department of Pharmacy, Seoul National University, Seoul, Korea). The MF was extracted with water (1.0 kg : 10 L) for 3 times, and filtered and concentrated to yield (167.32 g), and then suspended in distilled water and partitioned with hexane, EtOAc, and n-BuOH to give hexane-soluble (33.12 g), EtOAc-soluble (19.89 g), and n-BuOH-soluble (43.39 g) fractions, respectively. Among in the hexane-soluble obtained: fraction 1 (2.3 g), fraction 2 (1.6 g), fraction 3 (3.6 g), fraction 4 (2.9 g), fraction 5 (3.1 g), and fraction 6 (2.6 g), respectively. Finally, ursolic acid, ecdcysteron, and ergosterol peroxide were isolated as active principle compounds from fraction 6 by 5α-reductase inhibition (Fig. 1).
5α-reductase assay
The 5α-reductase assay was performed as described previously15 with slight modifications. Inhibitory activity of 5 α-reductase was determined by two special reactions; first, the replace extract with 0.2 ml of 50% ethanol to complete the reaction (rxn); second, an enzyme blank (ctrl) is added with 5.0 ml of dichloromethane before adding nicotinamide adenine dinucleotide phosphate. The peak area ratio (r) of testosterone/internal standard was used to calculate the percentage inhibition rate (%) by the following formula:

Animal experimental protocol
Five-week-old male C57BL/6J mice (n=6) were controlled temperature (23℃±2℃), humidity (50%±10%), and a 12-hour light-dark cycle, and then acclimatization for seven days. All mice were carefully removed from the dorsal areas (2×4 cm) with animal clipper containing wax-rosin mixture16 and MF extract was applied daily on the dorsal skin of mice on day 1 post depilation for 30 days. Animal study was conducted in accordance with the Hallym University protocol (Hallym-2012-70-1).
Histological analysis
The dorsal skin samples were fixed in 4% formalin and embedded in paraffin block, cut into 5 µm sections, and stained with hematoxylin and eosin to confirm hair follicle. Three different cross-sectional areas and the number of hair follicles were calculated using an image analysis program (Image-Pro Plus ver. 6.0; Media Cybernetics, Silver Spring, MD, USA).
Cell culture and proliferation
The human hair dermal palilla cells (HHDPCs) were incubated in mesenchymal stem cell medium (5% fetal bovine serum, 1% penicillin-streptomycin, and 1% mesangial stem cell growth supplement) at 37℃ in 5% CO2. HHDPCs were plated at 5×103 cells per well in 96-well plates, and incubated in the presence or absence of ursolic acid, ecdcysteron, and ergosterol peroxide. After 24 and 48 hours of culturing HHDPCs, added 20 µl of (3-(4, 5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium) solution to each well measured optical density at 550 nm using a microplate reader (Sensident Scan; Labsystems, Helsinki, Finland).
Reverse transcription-polymerase chain reaction (RT-PCR) and real-time polymerase chain reaction (PCR)
The total RNA was isolated from HHDPCs and dorsal dermal tissues and quantified using the NanoDrop-2000 (Thermo Fisher Scientific; Waltham, MA, USA). The cDNA synthesis was performed using a cDNA reverse transcription kit and real-time PCR was performed using the LightCycler real-time PCR System (Roche Applied Science, Indianapolis, IN, USA). The annealing temperatures and primers used for PCR reactions were listed in Table 1.
Western blot Analysis
The total protein (30 µg) from HHDPCs and dorsal dermal tissue was lysed in Lysis buffer (Thermo Fisher - Scientific) and separated by sodium dodecyl sulphate-polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride membranes. After blotting, membranes were probed with transforming growth factor-beta 1 (TGF-β1), Dickkopf-1 (DKK-1), insulin-like growth factor 1 (IGF-1), keratinocyte growth factor (KGF), and actin (Santa Cruz Biotechnology, Santa Cruz, CA, USA), and incubation with secondary antibody for 1 hour at romtemperature (25℃±1℃), immunoreactive proteins were visualized using an enhanced chemiluminescence substrate and quantified by densitometric analysis.
Statistical analysis
Student's unpaired t-test or one-way ANOVA were calculated with GraphPad Prism program (GraphPad Software Inc., San Diego, CA, USA). A p-value <0.05 were considered statistically significant. Results are represented as the mean±standard error, and multiple comparisons were carried.
RESULTS
MF extract on hair regeneration in C57BL/6J mice
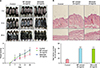
To investigate whether MF extract promotes hair growth, we induced the anagen phase of hair growth in C57BL/6J mice. After 30 days, MF extract induced telogen-to-anagen conversion earlier than vehicle treatment. During the experiment, visual scores were given weekly for hair growth (Fig. 3A). In the representative longitudinal section of histologic studies showed that MF extracts increased size of hair follicles and longer depth as compared with control group (Fig. 3B).
Effect of MF extract on growth factors in dorsal dermal tissues of C57BL/6J mice
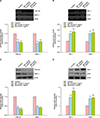
In this study, we measured the gene and protein expression levels of TGF-β1, DKK-1, IGF-1, KGF, and actin. As shown Fig. 4A and C, MF extract downregulated the expression of TGF-β1 and DKK-1 compared to vehicle treatment. However, it upregulated the gene expression of IGF-1 and KGF (Fig. 4B, D).
Isolation of active components of the MF extract
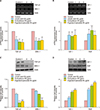
To achieve a satisfactory separation of the multiple components in the MF extract, the 5α-reductase activities of each fraction was examined using high-performance liquid chromatography (HPLC). The EtOH extract was fractionated into hexane, EtOAc, and n-BuOH extracts. The hexane extract significantly inhibited 5α-reductase inhibition activity by 62.2%, whereas EtOAc and n-BuOH extracts showed no effect (Fig. 5A). We obtained six fractions from the hexane extract, and their 5α-reductase inhibition activities were assessed. Fraction 6 of the hexane extract significantly inhibited 5α-reductase activity by 63% (data not shown). Furthermore, we identified ursolic acid, ecdcysteron, and ergosterol peroxide as the major active compounds in fraction 6. In addition, 5α-reductase activity was inhibited by ursolic acid, ecdcysteron, and ergosterol peroxide by 53.2%, 37.7%, and 35.0%, respectively (Fig. 5B).
Effect of ursolic acid, ecdcysteron, and ergosterol peroxide on the proliferation and gene expression in HHDPCs
To assess the action mechanism responsible for the hair regeneration effects of MF extract in C57BL/6J mice, we investigated the effects of its major compounds on proliferation and gene expression in HHDPCs. Minoxidil (100 µM) was used as a positive control. Proliferation of HHDPCs treated with MF extracts at 50 or 100 µg/ml for 72 hours was 121.8% and 119.0% higher, respectively, than that of the control group (Fig. 6A). Furthermore, ursolic acid, ecdcysteron, and ergosterol peroxide also improved the proliferation of HHDPCs in time- and dose-dependent manner (Fig. 6B). We also examined the expression of genes responsible for androgen signaling and hair cycle regulation in HHDPCs. As shown in Fig. 7A and C, the gene and protein expression TGF-β1 and DKK-1 were lower in cells treated with ursolic acid, ecdcysteron, and ergosterol peroxide than in control cells. However, these compounds induced the expression of IGF-1 and KGF, responsible for hair growth, in HDDPCs (Fig. 7B, D).
DISCUSSION
Hair is not only provides protective but also used to express personal beliefs or social position. Therefore, androgenic alopecia is considered a serious cosmetic problem in modern society. In our preliminary study, we screened 100 plants and found that MF extract showed the most effective 5α-reductase inhibitory activity as confirmed by HPLC (data not shown). Based on this previous result, we explored whether MF extract promotes the growth of HHDPCs and hair regeneration in C57BL/6J mice. In the present study, our findings suggest that MF extract contributes to hair growth-promoting effect and regulated the expression of growth factors and inhibited 5α-reductase activity.
The conversion of testosterone to dihydrotestosterone (DHT) by the enzyme 5α-reductase, and DHT causes hair loss917. Therefore, 5α-reductase is considered as one of the most important targets for the development of hair loss drugs. In addition, TGF-β1 and DKK-1 are important diffusion factors regulation the interaction between papilla-epithelium1819. The expression of the Wnt (winglesstype MMTV integration site family) ligand antagonist DKK-1 has been found to be up-regulated in response to DHT and reported to cause apoptosis in the bald scalp of patients with AGA2021. On the other hand, hair growth factor has also become one of the targets of hair loos treatment22. It is well known that IGF-1 and KGF is fundamental to stimulate the growth of hair follicles in HHDPCs2324. In the present study, MF extract inhibits 5α-reductase activity as well as downregulates TGF-β1 and DKK-1 gene and protein expression in C57BL/6J mice and stimulated increases in IGF-1 and KGF mRNA and protein levels in HHDPCs.
Based on these observations, MF extract represents a novel 5α-reductase inhibitor and has been selected for further research, including screening of active component. In this study, we identified and isolated active components of MF, such as ursolic acid, ecdcysteron, and ergosterol peroxide, guided by a 5α-reductase inhibition assay using HPLC. Among them, ursolic acid showed an inhibition of 53.2% at 50 µg/ml in accordance with previous results25; it was obtained from fraction 6 of MF extract (hexane fraction). Interestingly, the inhibitory activity of ursolic acid was not greater than that of the MF extract and other components. Thus, synergistic effect, two other major components isolated from the MF extract (hexane fraction), which are ecdcysteron and ergosterol peroxide, also showed lower inhibitory activity than that of the MF extract. Therefore, the three compounds have synergistic effects and are ubiquitous phenomena in natural products26. To understand their hair growth-promoting effects in HHDPCs, the androgen signaling pathway and key growth factor genes was studied. In in vivo study, we examined the expression of genes responsible for affecting the health of hair follicles and their growth in AGA. As shown in Fig. 7, the mRNA and protein expression of TGF-β1 and DKK-1 were decreased by ursolic acid, ecdcysteron, and ergosterol peroxide in HHDPCs when compared to control. However, these compounds markedly increased the expression of growth factors, such as IGF-1 and KGF, in HHDPCs. Therefore, our findings indicate that these bioactive compounds may be further studied for the treatment of AGA. Collectively, our results demonstrated that MF extract has a potent hair growth-promoting effect on C57BL/6J mice, suggests that MF extract and its active compounds may become a good candidate for promoting hair growth.




 XML Download
XML Download