

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Acute pancreatitis (AP) is a common and potentially lethal acute inflammatory disease associated with the release of digestive enzymes to the pancreatic interstitium and to systemic circulation, resulting in elevated production and release of cytokines, and ultimately in destructive local and systemic effects [1]. Systemic manifestations of the AP are mediated by a variety of pro- and anti-inflammatory mediators released from the pancreas and various other sources during the course of the disease. Local recruitment and activation of inflammatory cells in AP lead to the production of proinflammatory cytokines, such as interleukins (IL)-6, IL-8, and tumor necrosis factor-alpha (TNF-α), which are mostly studied as markers of severity of AP [2]. Activated pancreatic macrophages release TNF-α in response to local tissue damage, acting locally to aggravate AP and systemically with IL-6 to increase capillary permeability and promote leukocyte adherence and extravasation, leading to multiple organ failure [3]. Another commonly examined marker for AP is CRP [4].
Oxidative stress is known as the major pathogenic factor in the AP [5]. Cerulein, a cholecystokinin (CCK) analogue (pGlu-Gln-[Met3]-CCK8, sulphated), produces large amounts of reactive oxygen species (ROS) which are important mediators in the initiation and development of pancreatitis. ROS activate the oxidant-sensitive nuclear transcription factor (nuclear factor kappa B, NF-κB) and thus induce cytokine expression in the pancreatic acinar cells [6]. Among several animal models of experimental pancreatitis that exhibit the biochemical, histomorphological, and pathophysiological similarities to various aspects of human pancreatitis, cerulein pancreatitis was shown to be one of the best-characterized and widely used experimental models [7]. Different doses of cerulein causes variations in the pancreatic secretion of amylase and lipase resulting in pancreatitis [1], which is characterized by a dysregulation of the production and secretion of digestive enzymes, particularly, the inhibition of pancreatic secretion and an elevation in their serum levels, cytoplasmic vacuolization and the death of acinar cells, interstitial edema, and an infiltration of inflammatory cells into the pancreas [8].
Severe AP is associated with high morbidity and mortality due to the development of pancreatic and extrapancreatic necrosis, their subsequent infection and multisystem organ failure [9]. Based on the pathophysiology of AP and the basic science research conducted providing evidence for promising pharmacologic therapy, many clinical studies have been performed assessing the effectiveness of these therapies including anti-secretory agents, protease inhibitors, immunomodulators, anti-inflammatory agents, and antioxidants [1011]. Different anti-inflammatory agents and antioxidants such as ROS scavengers targeting the production of superoxide in the activated neutrophils, the expressions of the inflammatory cytokines, or the NF-κB activation, have been investigated in the treatment of AP [12]. However, there is still need to find the underlying mechanism of the disease that may serve as the potential therapeutic target in the development of new drugs, which may serve as a minimally invasive approach in the management of AP.
Ruscogenin ([1β, 3β, 25R]-Spirost-5-ene-1,3-diol), a main steroid sapogenin of a traditional Chinese plant called ‘Ophiopogon japonicus,’ was first isolated from ‘Ruscus aculeatus’ and has been found to have notable anti-inflammatory and antithrombotic activities [13]. Previous studies have revealed that the potent anti-inflammatory mechanism of ruscogenin is essentially related to the inhibition of nuclear factor kappa B (NF-κB) signal pathways and of the expression of intercellular adhesion molecule-1 (ICAM-1) [13]. However, there are no reports about the therapeutic effects of ruscogenin on AP, and the underlying mechanisms remain unclear. Thus, we aimed to evaluate the possible therapeutic effects of ruscogenin on the cerulein-induced AP by using macroscopic, histopathological, immunohistochemical, biochemical, and electron microscopic methods.
METHODS
Animals
All experimental procedures of this study were approved by the Research Ethics Committee of The Bagcilar Training and Research Hospital of Health Science University approved (protocol number: 2016/24, date of approval: 12 August 2016).
Thirty-two female Sprague-Dawley rats (250 ± 30 g), supplied by the Bagcilar Training and Research Hospital Animal Center (BADABEM), Istanbul, Turkey, were kept under laboratory conditions of the same center, housed in a controlled room with 12-hour light-dark cycles at 22℃, and fed with standard pellet chow including 21% protein and daily fresh water. Exclusion criteria were a weight loss of more than 20%, irregular nourishment and less drinking water, and a significant decrease in response to stimuli during the experiments.
All rats were separated in cages of 8 rats each and were divided into 4 random groups (n = 8) as follows;
Group 1. Sham group received 2 mL/kg/day amount of isotonic solution (IS) by gavage for 4 weeks.
Group 2. Control (AP) group was injected with a 20-μg/kg dose of cerulein 4 times at 1-hour intervals intraperitoneally.
Group 3. Prophylaxis group received a 3-mg/kg/day dose of ruscogenin (H10000934, Abdi İbrahim Drug Industry and Trade Inc., Istanbul, Turkey) daily for 4 weeks by gavage, following 4 intraperitoneal injections of 20-µg/kg cerulein at 1-hour intervals.
Group 4. Treatment group received a 20-µg/kg dose of cerulein 4 times at 1-hour intervals, simultaneously with a 3-mg/kg/day dose of ruscogenin applied by gavage.
Induction of AP
AP was induced by intraperitoneal administration of 20 µg/kg of cerulein 4 times at 1-hour intervals. Twelve hours after the last dose of cerulein, the rats were sacrificed by cardiac puncture under 80-mg/kg ketamine (Ketalar, Pfizer, Turkey) and 5-mg/kg xylazine (Rompun, Bayer, Turkey) anesthesia, and approximately 4- to 5-mL blood samples were collected during intracardiac puncture.
There were no anesthetic complications or death which may conduce to exclude any rat from the experimental procedures. There were no side effects like inflammation on wounds or vomiting due to oral medication.
Morphological analysis
After sacrificing the rats, the pancreatic tissue obtained was washed rapidly with saline and fixed for 72 hours in 10% neutral buffered formalin for light microscopic examination. Tissues were then dehydrated by passing through a rising alcohol series (70%, 90%, 96%, 100%), cleared by keeping in xylene for 2 × 10 minutes. Hematoxylin and eosin staining was applied to the sections obtained from paraffin blocks with a thickness of approximately 4 µm. All stained sections were evaluated on a light microscope (Olympus BX51, Olympus, Tokyo, Japan) with the camera attachment (Olympus DP72, Olympus) by 2 experienced histologists, who were unaware of the experimental groups. Pancreatic injury was evaluated (Table 1) based on an adapted version of a semiquantitative evaluation [14]. The hypertrophy, autophagy and necrosis of acinar cells, vascular damage, interstitial edema, and ductal variation were given a histopathological score as 0, none; 1, mild; 2, moderate; 3, severe.
Immunohistochemical analysis
In accordance with a previously described method [15], the paraffin sections were stained immunohistochemically by using streptavidin-biotin-peroxidase method with monoclonal and polyclonal antibody tagged to indicate NF-κB (anti-NF-κB p65 antibody, SAB4502610, Merck, Darmstadt, Germany). Biotinylated secondary antibodies and streptavidin-biotin-peroxidase conjugates, 3,3′-Diaminobenzidine were purchased from Invitrogen (HistostainPlus Kit, Life Technologies, Carlsbad, CA, USA). Counter staining was performed by Mayer Hematoxylin and positively staining with relevant antigens were analyzed semiquantitatively in terms of staining intensity (0, no staining; 1, weak reactivity; 2, moderate reactivity; 3, strong reactivity) [16].
Biochemical analysis
Blood and pancreatic tissue samples were collected for biochemical analysis. Blood cytokine levels of IL1β, IL-6, TNF-α, CRP, high-sensitivity CRP (hs-CRP) and the biochemical parameters (amylase, lipase) and ICAM-1 were examined by enzyme-linked immunosorbent assay (ELISA). The tissue amounts of antioxidant system-related parameters, nitric oxide (NO) and malondialdehyde (MDA) and ICAM-1 were also analyzed. Blood samples were allowed to clot at room temperature, centrifuged for 15 minutes at 1,000 ×g. The supernatants were collected. Pancreatic tissues were minced to small pieces and rinsed in ice-cold phosphate buffered saline to remove excess blood thoroughly. Tissue pieces were homogenized for 8 minutes at 20,000 rpm in 100 µL 0.02 M ethylenediaminetetraacetic acid. Homogenates were centrifuged for 5 minutes at 5,000 ×g and supernatants were collected.
ELISA methods were applied according to the manufacturer's instructions without any modifications by using rat sICAM/CD54 ELISA Kit (E-EL-R0369, Elabscience, Houston, TX, USA), CRP ELISA Kit (E-EL-R0022, Elabscience), hs-CRP ELISA kit (E-EL-R0506, Elabscience), IL-6 ELISA Kit (E-EL-R0015, Elabscience), IL-1β ELISA kit (E-EL-R05012, Elabscience), MDA ELISA kit (E-EL-0060, Elabscience), NOS1/nNOS ELISA kit (E-EL-R1438, Elabscience), TNF-α ELISA kit (E-EL-R0019, Elabscience), Amylase Alpha 1 (AMY1) ELISA kit (E-EL-R2544, Elabscience), endothelial lipase ELISA kit (E-EL-0365, Elabscience). Optical densities were read on a plate reader set at 450 ± 2 nm. The concentration of each parameter in the samples was calculated from the standard curve, multiplied by the dilution factor and was expressed as mean ± standard error of the mean (SEM).
Statistical methods
All data are expressed as means ± SEM with all rats per group. Instat statistical package (GraphPad Software, San Diego, CA, USA) was used. Following the assurance of normal distribution of data, 1-way analysis of variance with the Tukey-Kramer post hoc test was used for multiple comparison. Values of P < 0.05, P < 0.01 and P < 0.001 were regarded as significant.
RESULTS
Histopathological findings
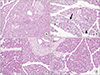
The sham group had a normal structure of exocrine pancreas including cells with normal basal nuclei and apical acidophilic cytoplasm, as well as a regular islet morphology in endocrine pancrease (Fig. 1A), whereas the cerulein-induced pancreatitis group (control group) had increased acinar cell hypertrophy, autophagy, necrosis, and vascular damage, interstitial edema, and ductal variation (Fig. 1B). The histopathological changes in the control group were statistically significant (P < 0.05) (Table 2). In addition, dilated interlobular spaces and scattered islet cells were found in the control group (Fig. 1B). On the other hand, the pancreatic tissues in prophylaxis and treatment group had a significant decrease in the hypertrophy, autophagy, necrosis of acinar cells, and vascular damage, interstitial edema, and ductal variation compared with the control group (Table 2). Moreover, the general appearance of Langerhans islets in both ruscogenin groups was observed as normal as in the sham group (Fig. 1C, D). There was no significant difference between the histopathologies of prophylaxis and treatment groups, as well as sham group.
Immunohistochemical findings
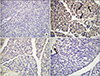
The number of NF-κB positive cells in the pancreatic tissues was very low in the sham group (Fig. 2A) while the number was highest in the cerulein-induced pancreatitis group, especially in the periacinar, perivascular, and periductal regions (Fig. 2B). The immunohistochemical scores showed a considerable increase in immunoreactivites of NF-κB in the control group compared with other groups (Fig. 3). On the other hand, pretreatment and posttreatment with ruscogenin led to a dramatic decrease in the number of positive cells (Fig. 2C); in fact, there were rare positive cells in the perivascular region of pancreases in the treatment group (Fig. 2D). The immunoreactivity scores for NF-κB were decreased to the normal levels both in the prophylaxis and treatment groups compared with the control group (P < 0.001) but there was no statistical difference between prophylaxis and treatment groups (Fig. 3).
Biochemical findings
The biochemical parameters obtained from blood by ELISA method are introduced in Table 3, and those from pancreatic tissues are in Table 4. As a well-known marker of the severity of AP in blood, CRP and hs-CRP levels were significantly increased in the pancreatitis (control) group compared with the sham group (P < 0.001). Ruscogenin treatment before and after cerulein-induced pancreatitis significantly reduced CRP and hs-CRP to normal levels but the reduction in hs-CRP was not as efficient as in CRP levels.
The analysis of ICAM-1 in blood, a cell surface glycoprotein which is typically expressed on endothelial cells and cells of the immune system, showed a significant increase in the pancreatitis (control) and prophylaxis groups compared with the sham group (P < 0.001), while reduced dramatically in the treatment group (P < 0.001). ICAM-1 protein levels in the pancreatic tissues increased considerably in the control and prophylaxis groups compared with the sham group (P < 0.001 and P < 0.01, respectively), while ruscogenin treatment after pancreatitis induction reduced ICAM-1 levels dramatically, even falling under the levels of sham group (P < 0.001 vs. control group).
Blood IL-6 amounts, one of the inflammatory markers, in the pancreatitis groups were greater than the sham group; the significance was higher in the control group (P < 0.001) than the treatment groups (P < 0.001). Another inflammatory marker, IL-1β, was highly elevated in blood of the control group compared to the sham group, with a marked significance (P < 0.001), while the ruscogenin-treated rats had a lowered amount of IL-6 and IL-1β but failed to decrease to the levels of sham group (P < 0.001 vs. sham group). Pretreatment with ruscogenin was more successful than posttreatment to decrease the serum IL-6 and IL-1β amounts, but the difference was not significant.
The amount of another inflammatory marker, TNF-α, was elevated in blood of the control group and even in the ruscogenin-treated groups with a significant difference compared to the sham group (P < 0.001) though there was still a difference between the control and ruscogenin-treated rats (P < 0.001 vs. control group). However, there was no difference between the prophylaxis and treatment groups.
The amount of one of the pancreatic enzymes, amylase, markedly increased in the pancreatic control group compared with the sham group (P < 0.001). The prophylaxis group showed a significant decrease in amylase levels compared to the control group (P < 0.001) while the decrease in the treatment group was not statistically different compared with the control group. The other enzyme, lipase, increased dramatically in the control and even in the ruscogenin treatment groups (P < 0.001), with the difference between control and ruscogenin-treated groups also statistically significant (P < 0.001). Ruscogenin successfully reduced the amylase and lipase levels for both pre- and posttreatment.
Measuring the amount of MDA, one of the toxic stable decomposition products in the pancreatic tissue, the control, prophylaxis, and treatment groups showed significant elevations in comparison to the sham group (P < 0.001). Both pre- or posttreatment with ruscogenin failed to reduce MDA levels in the pancreatic tissues (Table 4).
One of the constitutive NO synthase enzymes, nitric oxide synthase 1/neuronal nitric oxide synthase (NOS1/nNOS), showed a dramatic increase in the tissues of pancreatitis control group, that was more than 5 times the amount measured in the sham group (P < 0.001). Ruscogenin treatment reduced the amount of NOS1/nNOS significantly in the prophylaxis and treatment groups, compared with the control group (P < 0.001). There was no considerable difference between the prophylaxis and treatment group.
Ultrastructural findings
TEM micrographs of endocrine cells in the sham group showed regular structures of heterochromatic nuclei, evenly distributed rough endoplasmic reticulum (RER), and mitochondria surrounded with normal capillaries (Fig. 4). Exocrine pancreas of the sham group had a number of regular zymogen granules in the cytoplasm, intact mitochondria, euchromatic oval nuclei, and RER in normal ultrastructure. However, the endocrine cells of AP group had a large number of vacuoles with different ingredients and in varied sizes, as well as disrupted mitochondria with reduced cristae, and degenerated RER with dilated cisterna. The acinar cells of this group had a reduced number of zymogen secretory granules, mitochondria with varied shape of cristae and many vacuoles in the cytoplasm (Fig. 4). The endocrine cells of prophylaxis group had also few vacuoles located in peripheral cytoplasm, but regular intact mitochondria, and evenly distributed RER. The acinar cells of prophylaxis group had also normal and evenly distributed mitochondria and RER, a large number of zymogen granules, as well as a few small vacuoles (Fig. 4). The endocrine cells of treatment group showed regular structures as in the sham group, except a few slightly degenerated mitochondria. The acinar cells of treatment group also had RER with slightly dilated cisternae, a considerable number of smaller zymogen granules with enlarged peripheral spaces, intact normal mitochondria, but no vacuoles (Fig. 4). In brief, the ruscogenin treatment ameliorated the cerulein-induced ultrastructural degenerations in the endocrine and exocrine cells of the pancreatic tissue, especially in the posttreatment group.
DISCUSSION
This study investigated the protective and therapeutic effects of ruscogenin, a major steroidal sapogenin of Radix Ophiopogon japonicas, in an AP rat model. We demonstrated the molecular mechanism of its protective and therapeutic actions accomplished by improvements in histopathology, by decreasing blood cytokine levels of CRP, hs-CRP levels, TNF-α, IL-6, IL-1β, ICAM-1, by reducing the pancreatic enzymes amylase and lipase in blood, and by suppressing the expression of NF-κB, ICAM-1 and NOS1 in pancreatic tissues. The ruscogenin treatment also improved the cerulein-induced ultrastructural degenerations in the endocrine and exocrine cells of the pancreatic tissue, especially in the treatment group.
Many findings of AP in humans are comparable and able to be reproduced in the cerulein-induced AP models, including hyperamylasemia and diverse histopathological findings: infiltration of inflammatory cells within the pancreas, pancreatic edema, acinar cell vacuolization, and the presence of activated pancreatic enzyme within the pancreas. Apart from injury to the acinar cells, ductal and endocrine cells are not damaged [17]. In the present study, we successfully showed the destructive effect of cerulein-induced AP in our model, as elevated amounts of amylase and lipase in blood, and deteriorating histopathology of the pancreas as increased hypertrophy, autophagy, necrosis and vacuolization of acinar cells, and vascular damage, interstitial edema, and ductal variation, as well as the ultrastructure of the endocrine and exocrine cells.
Natural polyphenols have been demonstrated to have several sanative roles in the gastrointestinal organs. Among these roles, antioxidant and anti-inflammatory properties, are the most striking and remarkable ones for the researchers [11]. Moreover, the protective and therapeutic effects of various traditional and supplementary drugs on AP are related to their polyphenol contents [18]. The extracts of the R. aculeatus plant have been used in a large number of studies in terms of their pharmacological features. Two main active ingredients of this plant responsible from its pharmacological effects, ruscogenin and neoruscogenin recently gain attention for their anti-inflammatory and antioxidant characteristics, as well as their vasoconstrictive and venotonic features. In our previous study, we have also demonstrated that the ruscogenin treatment reduced ulceration in gastric tissues in a chronic ulcer model established by acetic acid induction [19]. The healing effects of ruscogenin in acetic acid-induced gastric ulcer may be due to inhibiting oxidative stress, promoting antioxidant mechanisms and the inhibition of lipid peroxidation by maintaining a balance in collagen content of extracellular cell matrix and ultrastructure [19]. In the present study, we discovered the healing and protective effects of ruscogenin in a cerulein-induced AP rat model by histopathological, immunohistochemical, biochemical, and electron microscopical analysis.
In our previous study, ruscogenin was considered to exert its potent efficacy in gastric ulcer via anti-inflammatory actions through reducing TNF-α, IL-6 and IL-8, antioxidant actions through enhancing glutathione activities, in addition to depression in oxidative stress levels and suppressive effects on lipid metabolism [19]. In the present study, the efficient protective and therapeutic effects are probably due to anti-inflammatory actions through decreasing blood cytokine levels of CRP, hs-CRP levels, TNF-α, IL-6, IL-1β, ICAM-1, by reducing the pancreatic enzymes amylase and lipase in the blood, and by suppressing the expression of transcription factor NF-κB, and by staving off the augmentation in NO production through suppressing NOS1 in pancreatic tissues.
The pathogenesis of AP is not clear. Recently, studies have focused on trypsinogen activation, pancreatic microcirculation malfunction, calcium overload, and inflammatory and antioxidant pathways [20]. Natural anti-inflammatory agents, such as resveratrol and curcumin, demonstrate a modest preventative role in the treatment of AP [1821]. However, specific and effective therapies and protective mechanisms are lacking. Significant therapeutic effect of resveratrol was noticed in rats with AP by inducing apoptosis of pancreatic acinar cell as a consequence of upregulated FasL gene expression [22]. The mechanism responsible for this protective activity is based on the upregulation of antiapoptotic Bcl-2 and downregulation of proapoptotic Bax and caspase-3 [18]. It seems that resveratrol has a selective bioactivity, being proapoptotic on acinar cells and antiapoptotic on other types of cells (e.g., hepatocytes). The resveratrol, with its metabolite (dihydroresveratrol), exerted a dose-dependent anti-inflammatory effect through NF-κB inhibition and decreased expression of TNF-α, IL-1β, IL-6, and IL-8 in the pancreas in various animal models of pancreatitis (e.g., cerulein) [23]. Therefore, we investigated the expression of NF-κB in pancreatic tissue and the cytokine levels (TNF-α, IL-1β, IL-6) biochemically in our AP model and found that the ruscogenin treatment inhibited their biosynthesis markedly in serum of rats, consistent with the notion that the inhibition of inflammatory cytokines is a therapeutic mechanism against pancreatic damage.
NO is a key signaling molecule in the pathogenesis of inflammation, but its role in AP has not been enlightened clearly yet [24]. There are contradictory findings concerning the effects of polyphenols on the production of NO and NOS. For example, resveratrol was considered to activate endothelial NOS which was found to be protective against cerulein-induced AP in mice, while inducible NOS exhibited proinflammatory effects in the same animal model [25]. Therapeutic blockade of inducible NOS (iNOS) or nNOS provides benefits in terms of inhibition of the acute pancreatitis-related abdominal hyperalgesia, while iNOS inhibition also ameliorates inflammatory cell influx to the pancreas and reduces the resultant hyperamylasemia and NOx levels, thus representing alternative pharmacological strategies for treatment of clinical severe pancreatitis. nNOS staining was reported (without NADPH-d analysis) in rat acini with agonist-evoked [Ca2+] oscillations [26], but nNOS has not been shown in cerulein-induced AP in rats before. In the present study, NOS1/nNOS showed a dramatic increase in the tissues of AP rats, while ruscogenin pre- and posttreatment reduced the amount of NOS1/nNOS significantly, showing a protective and therapeutic effects of ruscogenin via altering NO pathways.
As a limitation in the present study, we did not examine the expressions of other 2 isoforms of NOS, namely endothelial NOS (eNOS) and iNOS. eNOS is the main isoform influencing the initiation of cerulein-induced AP and eNOS-derived NO exerts a protective effect through actions on nonacinar cell types, most likely endothelial cells [27]. However, all aspects of the biochemical analysis for nNOS reveals the effectiveness of ruscogenin in AP through inhibiting nNOS production and thus, the valuable information added to the literature by this study is incontrovertible.
Polyphenols have been implicated to induce pancreatic superoxide dismutase (SOD) and inhibit the production of MDA, a marker of lipid peroxidation, within the first 6 hours of AP induction in rats [22]. In our study, we were able to show an inductive effect of cerulein to increase lipid peroxidation in pancreatic tissues, indicated by increasing MDA levels. However, ruscogenin failed to decrease this effect, probably due to long-term treatment. However, a study investigating the chemopreventive potential of Tribulus terrestris, a tropical plant which contains ruscogenin, in rats with skin carcinogenesis, found that treatment of the extract increase reduced glutathione levels and decrease MDA formation. The decreased lipid peroxidation in the liver homogenate of Tribulus treated animals is correlated well with the induction of antioxidant enzymes above basal level [28]. A single-blind, randomized, placebo-controlled study evaluated the effects of oral mixture of curcumin with piperine (5 mg) in tropical pancreatitis, a type of chronic pancreatitis. The herbal formulation significantly reduced the erythrocyte MDA level [29]. On the contrary, pre- or posttreatment with ruscogenin failed to reduce MDA levels in the pancreatic tissues. Since lipid peroxidation has been implicated in the pancreatitis, it is assumed that antioxidant components of the plant extracts could act as potential drugs to prevent or cure the disease [2223]. To verify this hypothesis, in future, SOD, myeloperoxidase, or glutathione amounts in pancreatic tissues and blood of rats should be investigated to show the exact antioxidant effects of ruscogenin in AP.
Ruscogenin has been found to have valuable anti-inflammatory and antithrombotic activities in inflammatory diseases. A dose-dependent anti-inflammatory effect of the crude steroidal saponin from the rhizomes of Ruscus aculeatus L. (Ruscaceae) was shown in 2 rat models of acute inflammation [13]. Another study suggested that ruscogenin considerably inhibited adhesion of leukocytes to a human umbilical vein endothelial cell line (ECV304) injured by TNF-alpha in a dose-dependent manner [30]. Huang et al. [13] reported that ruscogenin dramatically suppressed zymosan A-evoked peritoneal total leukocyte migration in mice in a dose-dependent manner, and also inhibited TNF-α-induced over expression of ICAM-1 both at the mRNA and protein levels and considerably suppressed NF-κB activation. Cao et al. [27] reported the protective effect of ruscogenin after ischemic stroke, by inhibiting IL-1β and Caspase-1, thus decreasing inflammation in vivo and in vitro. Since the main cause of AP is acute inflammation, we expected and observed an inhibitory effect of ruscogenin on NF-κB and ICAM-1 in cerulein-induced pancreatic tissues, and serum TNF-α, IL-6 and IL-1β, ICAM-1 levels, suggesting a possible molecular mechanism of ruscogenin to be anti-inflammatory effect through NF-κB, ICAM-1, TNF-α, IL-6 and IL-1β.
Accumulating evidence has demonstrated that traditional Chinese medicine reduces the levels of serum and urinary amylase, decreases the permeability of capillaries, depresses the production of inflammatory cytokines, inhibits neutrophilic granulocyte activation, and attenuates pancreatic injuries. These benefits block multiple steps in the development of AP according to experimental and clinical studies. Based on its characteristics of improved symptoms, reduced medical costs, and increased satisfaction of acute pancreatic patients, these medical agents appear to be a promising complementary and alternative therapy for the treatment of disease [2930]. In the present study, pre- and posttreatment by ruscogenin successfully attenuated pancreatic injury by reducing levels of the pancreatic enzymes amylase and lipase in the cerulein-induced AP rat model, as reflected by improved histopathologic scores.
In our previous study, we showed acetic acid-induced gastritis resulting in devastating damage on ultrastructure of chief and parietal cells while ruscogenin almost completely ameliorated these effects and preserved the ultrastructure, suggesting a correlation between cellular integrity and anti-inflammatory effects [19]. In this study, the ruscogenin treatment ameliorated the cerulein-induced ultrastructural degenerations in the endocrine and exocrine cells of pancreatic tissue, especially in the treatment group.
In conclusion, the present findings have demonstrated that the beneficial protective and therapeutic effects of ruscogenin appears to be mediated by diminished expression of NF-κB and ICAM-1 in the pancreatic tissue, and by reducing serum TNF-α, IL-6 and IL-1β, CRP, hs-CRP, amylase, lipase, and ICAM-1. Evidently, this nominates ruscogenin as a highly promising supplementary agent to be considered in the treatment of AP, and even as a protective agent against damages induced by disease.





 XML Download
XML Download