

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Staphylococcus aureus is an important zoonotic pathogen that can cause various diseases including bovine mastitis and endometritis in veterinary clinics, which seriously endangers the sound development of dairy cow farming [12]. Once successfully infecting the mammary gland, S. aureus will rapidly propagate, release a variety of toxins and extracellular enzymes, and form a biofilm, leading to increased resistance to antibiotics [34]. In recent years, the emergence of methicillin- and vancomycin-resistant strains, and especially their persistent propagation across animals and humans have seriously threatened public health [5]. Therefore, bovine mastitis and endometritis caused by S. aureus have become the most costly and hardest to prevent and control diseases in veterinary clinics.
As an important foodborne bacterium, S. aureus can release a variety of virulence factors including staphylococcal enterotoxins (SEs), hemolysin, and toxic shock syndrome toxin 1 (TSST-1) during propagation [67]. As a result, S. aureus and SEs contained in bovine milk are important factors causing food poisoning in human populations. Poisoning cases due to drinking bovine milk account for as high as 33%–50% of total food poisonings caused by bacteria and about 95% of staphylococcal food poisoning outbreaks are caused by the conventional SE genes sea to see [8910].
Xinjiang is one of China's most important milk-producing areas, with milk production in that region ranking as number one in western China. However, mastitis and endometritis in dairy cows have not only led to serious economic losses in dairy industry but also posed a great threat to food safety. Due to long-term use or abuse of antibiotics in dairy farms, drug-resistant strains have noticeably emerged in recent years. However, the phenotypic and molecular characteristics of S. aureus isolates from dairy cows in Xinjiang have not been completely described. The main purpose of the present study was to characterize the biofilm, antibiotics resistance, enterotoxin, and virulence profiles of S. aureus isolates from dairy cows in Xinjiang Province, China. Such information can provide deep insights into the biological characteristics of the endemic strains of this foodborne bacterium and contribute to food safety control.
MATERIALS AND METHODS
Isolation and identification of S. aureus
A total of 386 clinical samples were collected during 2015 and 2018 from bovine milk produced from 15 large-scale dairy farms in Xinjiang, China. Among them, 276 samples were collected from cows with mastitis and 110 from cows with endometritis. The collected samples were inoculated on Baird-Parker medium (Sigma, USA) at 37°C for 12–24 h. Then, suspected bacterial colonies with typical staphylococci characteristics were stained using the Gram method in order to inspect bacterial morphology. Colonies characterized as Gram-positive cocci were further subjected to catalase and coagulase testing. After DNA was extracted from the Gram-positive colonies of bacteria, the 16S rRNA gene was specifically amplified by polymerase chain reaction (PCR) for molecular level-based identification of the isolate [11].
Biofilm formation ability assay
The biofilm formation ability of the S. aureus isolates was semi-quantitatively determined using a microtiter plate-based adhesion (MPA) assay and the crystal violet staining method. According to their biofilm formation ability, the isolates were classified into four categories: 1) no biofilm formation ability (OD ≤ ODC, BF−); 2) weak biofilm formation (ODC < OD ≤ 2× ODC, BF+); 3) medium biofilm formation (2× ODC < OD ≤ 4× ODC, BF2+); and 4) strong biofilm formation (4× ODC < OD, BF3+) [1213].
Detection of biofilm formation-related genes and their transcription levels
The S. aureus isolates were inoculated into BHI medium (Sangon, China) and cultured overnight at 37°C. Genomic DNA was extracted using commercial kits (Qiagen, Germany) and used as templates for PCR amplification of genes clfA, clfB, fnbA, fnbB, fib, can, icaA, icaC, icaD, and other biofilm formation-related genes. Primers used for PCR are listed in Supplementary Table 1. In addition, the transcription levels of these genes were determined by using quantitative reverse transcription-PCR with the 16S rRNA gene as the internal control. Briefly, total RNA was extracted using Trizol (Invitrogen, USA) and reverse transcribed into cDNA using the PrimeScriptTM RT reagent kit (Takara, Japan). The obtained cDNA was used as a template in quantitative real-time PCR to analyze the transcription levels of genes related to biofilm formation using the FastStart Universal SYBR Green Master (ROX) kit (Roche, Switzerland). Each sample was measured in triplicate and data were calculated using the modified 2-ΔΔCt method [14].
Detection of enterotoxin genes
S. aureus genomic DNA was extracted using a bacterial genomic DNA extraction kit (Qiagen, Germany). Primers specific to the enterotoxin sea–see and seg–seo genes were designed and used in PCR to detect the sea–see and seg–seo genes (Supplementary Table 2) [15].
Drug susceptibility test
Briefly, the drug susceptibility of the S. aureus isolates to penicillin (10 U), oxacillin (1 μg), cefoxitin (30 μg), erythromycin (15 μg), clindamycin (2 μg), trimethoprim (5 μg), ciprofloxacin (5 μg), levofloxacin (5 μg), tetracycline (30 μg), chloramphenicol (30 μg), gentamicin (10 μg), rifampicin (5 μg), teicoplanin (30 μg), nitrofurantoin (300 μg), and fosfomycin (200 μg) (Sigma) were measured using K-B method and following the procedure described by the American Clinical Laboratory Standardization Institute (CLSI, 2015) [1016].
Determination of hemolytic activity
In brief, S. aureus isolates with different biofilm formation abilities were cultured overnight in tryptic soy broth (TSB) (Sangon, China) and diluted 109 times. A total of 100 μL of the diluted cultures were coated onto agar plates prepared using 100 mL TSB supplemented with 5 mL fresh sheep serum and 2 g agar and then incubated at 37°C for 24 h. The diameters of the hemolytic rings formed by each isolate were measured and the average diameter was used to evaluate the hemolytic activity of different S. aureus isolates [4].
Determination of half lethal dose (LD50)
A total of 8 S. aureus isolates with different biofilm formation abilities were prepared as 105/mL, 106/mL, 107/mL, 108/mL, and 109/mL suspensions of each isolate. Two hundred Kunming mice weighing 18–22 g were randomly assigned to 8 groups with 25 mice in each group. The five different suspensions of each isolate were injected intraperitoneally into 5 groups of mice with 5 repeats per suspension. The survival rate of mice in each group was observed and the LD50 of each isolate to Kunming mice was calculated using the modified Karber's method as reported previously [1718].
Bacterial load and histopathological analysis
Briefly, the mice were infected with a BF− S. aureus isolate, a BF3+ S. aureus isolate, or sterile PBS (control group). After infection, liver, spleen, lung, kidney, and small intestine samples were taken from each mouse under aseptic conditions. A small piece of the individual organs of mice in the same group was placed in a 1.5 mL sterile Eppendorf tube for measuring the organ bacterial load. The remaining organ blocks were fixed with 4% formaldehyde solution, prepared as 4 μm thick sections, stained with hematoxylin and eosin, and observed under a microscope to visualize histopathological changes [17].
RESULTS
Assay of biofilm formation ability of S. aureus isolates
A total of 164 S. aureus isolates were obtained from 386 bovine samples. Most of the isolates formed a biofilm in 96-well plates (Fig. 1A). The results of the MPA assays confirmed that 23 (14.0%) of the isolates did not form a biofilm (BF−), while 141 (86.0%) could form a biofilm (BF-positive). Of the 141 BF-positive isolates, 69 (42.1%) were BF+, 38 (23.2%) were BF2+, and 34 (20.7%) were BF3+ (Fig. 1B and C).
 | Fig. 1Determination of BF formation ability of S. aureus isolates. (A) Straining of BF in 96-well microtiter plates; (B) determination of OD600nm of BF in S. aureus isolates with different BF formation ability; (C) Number of isolates with different BF formation ability.BF, biofilm; OD, optical density; S. aureus, Staphylococcus aureus.
|
Measurement of genes related to biofilm formation
Among the BF-positive S. aureus isolates, the detection rate of the icaD gene was the highest (100%), and those of clfB and fib were the lowest (58.8%). By comparison, among the BF− isolates, the detection rate of the icaD gene was the highest (100%), and those of clfB and icaC were the lowest (65.2%) (Fig. 2A). Overall, the detection rates for icaA and icaD were higher than that of the other biofilm formation-related genes; whereas, the detection rates for the clfB, fnbA, and fib genes were highest in BF− isolates (Fig. 2B and Supplementary Fig. 1).
 | Fig. 2Determination of BF formation-related genes and their transcriptional levels in S. aureus isolates with different BF forming ability. (A) BF formation-related genes in BF-positive isolates of S. aureus; (B) Comparison of detection rate of BF formation-related genes in BF-positive isolates of S. aureus; (C) Relative transcriptional levels of BF formation-related genes.BF, biofilm; S. aureus, Staphylococcus aureus.
|
Transcription levels of biofilm formation-related genes in S. aureus isolates
The transcription levels of 9 genes related to biofilm formation were significantly different between the BF-negative and BF-positive isolates (p < 0.05). Among them, the transcription levels of 6 genes (fnbA, fnbB, cna, fib, icaA, and icaC) were significantly higher in the BF3+ isolates than those in the other isolates (p < 0.05) (Fig. 2B and C).
Detection of enterotoxin genes in S. aureus isolates
The detection rate for all enterotoxin genes (sea–seo) in the 164 S. aureus isolates was 96.3%. Among them, the detection rate of the novel enterotoxin gene seg was the highest (81.1%). The detection rate of the conventional enterotoxin genes (sea–see) in the 164 S. aureus isolates was 62.8%, of which, that of the see gene was the highest (57.3%). In addition, the detection rate of the enterotoxin genes was not correlated with the biofilm formation ability of the S. aureus isolates (Fig. 3 and Supplementary Fig. 2).

Determination of drug susceptibility of S. aureus isolates
Of 164 isolates of S. aureus, most isolates (81.7%) were resistant to penicillin, followed by trimethoprim (69.5%), cefoxitin (52.4%) and clindamycin (51.2%). In contrast, these isolates had low drug resistance to fosfomycin (6.1%), teicoplanin (10.4%), nitrofurantoin (14.6%), and rifampicin (26.8%). Furthermore, BF-positive isolates were resistant to cefoxitin (56.7%), erythromycin (48.2%), clindamycin (52.4%), chloramphenicol (31.2%), gentamicin (32.6%), rifampin (27.6%), and fosfomycin (7.1%), and those resistance levels were significantly higher than those of BF-negative isolates (26.1%, 34.7%, 43.4%, 26.1%, 26.1%, 21.7% and 0%, respectively) (p < 0.05) (Fig. 4).

Assay of hemolytic activity of the S. aureus isolates
All BF−, BF+, and BF2+ isolates produced obvious hemolytic rings. The BF− strains had the largest average hemolytic ring diameter (up to 13.2 mm), while the BF3+ strains had the smallest average hemolytic ring diameter of (up to) 1.6 mm. Moreover, isolates with a higher biofilm formation ability had weaker hemolytic activity (Fig. 5).
 | Fig. 5Determination of the hemolytic activity of S. aureus isolates with different BF formation abilities cultured on SBA. (A) Hemolysis of S. aureus on SBA; (B) Diameter of erythrocyte hemolysis ring.BF, biofilm; SBA, sheep blood agar; S. aureus, Staphylococcus aureus.
* indicates p < 0.05, *** indicates p < 0.001.
|
LD50 of S. aureus isolates with different biofilm formation abilities
Among the four groups of S. aureus isolates with different biofilm formation abilities (BF−: XJC-45 and XJC-91; BF+: XJC-A2 and XJC-73; BF2+: XJC-A1 and XJC-90; BF3+: XJC-3 and XJC-49), the LD50 levels differed significantly between the BF3+ and BF2+ isolates, as well as between the BF+ and BF− isolates (p < 0.05). The LD50 values of BF3+ isolates were markedly high, indicating they were less pathogenic to Kunming mice (Fig. 6).
 | Fig. 6Mortality and LD50 of mice with different BF formation ability infected with S. aureus strains. (A) LD50 of S. aureus isolates in Kunming mice; (B) Survival curves of mice infected via intraperitoneal injection with BF− and BF3+ isolates.BF, biofilm; LD50, half lethal dose; S. aureus, Staphylococcus aureus.
|
Histopathological changes in mice infected by S. aureus with different BF formation abilities
Pathological changes in mice infected by BF− isolates were more severe than those in mice infected by BF3+ isolates (Supplementary Fig. 3). The histopathological changes in liver sections revealed granulation and vesicle degeneration in hepatocytes, blurry boundaries of hepatic lobules, and dilated central veins. The histopathological changes in spleen included unclear boundaries between the red pulp and white pulp, red blood cell exudation in the red pulp, and spleen body atrophy. Pulmonary interstitial edema and hemorrhage, plasma cells, and lymphocyte infiltration were observed in lung tissue. The histopathological change in kidneys included renal interstitial vasodilatation and renal tubular epithelial cell degeneration (Fig. 7).
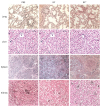
Bacterial load of mice infected by S. aureus isolates with different biofilm formation abilities
The invasiveness of BF− isolates to liver, spleen, lungs, kidneys, and small intestine of mice was stronger than that of BF3+ isolates (p < 0.05). The bacterial load was the highest in kidneys (range, 3.73 ± 2.36 to 3.50 ± 1.84) and the lowest in liver (range, 1.15 ± 0.21 to 0.99 ± 0.23) (Table 1).
Table 1
Comparison of bacterial loads in various organs between mice infected by BF3+ and BF− isolates of Staphylococcus aureus

![]()
DISCUSSION
In this study, we characterized the biofilm, antimicrobial resistance, enterotoxin, and virulence profiles of S. aureus isolates from dairy cows with clinical mastitis and endometritis. We obtained 164 S. aureus isolates from dairy cows in Xinjiang, China. Among them, 86.0% could form a biofilm, which was higher than those of S. aureus in other studies (58.0% to 69.8%) [161920], indicating that S. aureus isolates from dairy cows commonly possess biofilm formation ability and are mainly BF-positive.
Many studies have demonstrated that S. aureus can attach to biotic or abiotic surfaces to form a special structure, biofilm, which is an aggregated community of cells and is involved in adaptation to stressful environments [1421222324]. However, this structure can make S. aureus resistant to antibiotics or allow it to escape the host's immune system via restricting antibiotic penetration, limiting nutrients, and forming special phenotypes [212526]. The examination of biofilm formation ability showed that BF-positive strains were the predominant strains in dairy cows in Xinjiang, China, which may be related to the irrational use of antibiotics in dairy farms.
Previous studies have shown that several genes are involved in the biofilm formation process [1417222327282930]. PCR analyses in this study showed that 9 genes related to biofilm formation were detected in most isolates, and each isolate contained at least 4 genes related to biofilm formation; moreover, the number of genes detected was related to the biofilm formation ability of the isolate. Among the 9 genes related to biofilm formation, 6 genes encoding microbial surface components recognizing adhesive matrix molecules (MSCRAMM) had high detection rates (range, 61.5% to 82.9%). Moreover, the detection rates of the clfB, fnbA, and fibB genes were the highest in the BF− isolates, and relatively low in the BF2+ and BF3+ isolates. In addition, the transcription levels of the fnbA and fnbB genes were significantly high in BF3+ isolates, which was consistent with results reported in previous studies [3132], further confirming that fnbA and fnbB have important roles in biofilm formation in S. aureus strains.
Compared to bacteria in the planktonic state, bacteria in the biofilm state are significantly more resistant to antibiotics. Herein, we showed that BF-positive isolates had significantly greater resistance to erythromycin, clindamycin, ciprofloxacin, rifampicin, and fosfomycin than BF-negative isolates, and they appeared to be multidrug resistant, suggesting that biofilm formation ability of S. aureus is closely related to its drug resistance.
SEs are exogenous superantigens produced by S. aureus that can activate host systems to produce large amounts of pro-inflammatory cytokines and chemokines, which can result in fever, hypotension, and shock [15]. Therefore, they are key substances to be monitored in food production. Nazari et al. suggested that the distribution of SE genes in S. aureus isolates from different regions have obvious regional characteristics [11]. It is notable that the detection rate among all enterotoxin genes (sea–seo) was 96.3% in this study. Furthermore, of the examined genes encoding SEs, detection rates of the novel enterotoxin genes seg, sej, and sek were higher than those of the conventional ones and were obviously different from those reported by Wang et al. [33]. Accordingly, the high level of presence of novel enterotoxin genes is deemed an important molecular characteristic of the S. aureus isolates from dairy cows in Xinjiang. Therefore, surveillance of novel enterotoxins genes has the same importance as biofilm monitoring. Nevertheless, the pathogenicity and epidemiological risks associated with those isolates carrying novel enterotoxin genes need to be further explored.
Under stress, S. aureus can form a biofilm to resist the hosts’ immune responses and can re-infect the host in the planktonic state under favorable conditions, producing a persistent S. aureus infection [343536]. Therefore, exploring the intrinsic correlation between biofilm formation ability and pathogenicity in S. aureus strains is significantly important. Herein, we compared the LD50 and hemolytic activity of 8 S. aureus isolates with different virulence and biofilm formation ability and determined that isolates with a higher biofilm formation ability had weaker virulence, confirming the biofilm and virulence diversity character of S. aureus [37]. This phenomenon is possibly related to the influence of its ability to form biofilm on the release of virulence factors during the planktonic state [383940].
In summary, we, for the first time, have characterized the biofilm, enterotoxin, and virulence profiles of S. aureus isolates from dairy cows in Xinjiang, China. The present study demonstrated that BF-positive isolates were predominant; moreover, the BF-positive isolates had higher resistance but lower virulence than that of the BF-negative isolates. Considering that S. aureus is an important foodborne bacterium, which can be transmitted between dairy cows and humans, the high frequency of BF ability and the presence of novel enterotoxin genes in Holstein dairy cows in Xinjiang large-scale dairy farms may pose potential threats to food safety.
 XML Download
XML Download