

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Porcine epidemic diarrhea (PED) is one of the most catastrophic swine diseases and is characterized by severe diarrhea, vomiting, dehydration and even death [1]. The mortality rate of PED is up to 80-100%, especially in neonatal piglets [23], resulting in large economic losses for swine farms across the globe, including Europe, Asia and the USA for the past three decades [145], and making the development of an effective PED vaccine a necessity.
The currently used PED vaccines are live attenuated vaccines (LAV) or inactivated vaccines, which have inherent problems related to safety and cost-effectiveness [126]. In particular, the effectiveness of some of these vaccines has become controversial after an outbreak of PED in the United States [1], which consequently stressed the need for the development of a novel PED vaccine such as subunit vaccine. Subunit vaccines involve the use of an antigenic component of pathogenic bacteria or viruses that could be used as a vaccine instead of the entire pathogen. Despite relatively low immunogenicity, subunit vaccines have many advantages over traditional LAVs, including safety, simplicity and the ability to be mass produced through E. coli systems [7]. In particular, the cost-effectiveness of subunit vaccines is an important advantage in association with the economic objectives of the livestock industry. However, candidate antigens of the PED subunit vaccine, such as S0, are expressed as insoluble aggregates in E. coli that are not suitable for use as a vaccine [89]. Although S0 possesses enriched neutralization epitopes with a potential in cross-reactivity to various PEDV strains [10]. To overcome the low solubility and low immunogenicity of the PED subunit vaccine, the introduction of a molecular fusion adjuvant, such as flagellin protein, would be necessary.
Flagellin, a monomeric subunit of bacterial flagella, is one of many pathogen-associated molecular pattern (PAMP) proteins that are recognized by two germline-encoded receptors, TLR5 and NLRC4/NAIP5, on a number of innate and adaptive immune cells. Flagellin has been frequently used as a molecular fusion adjuvant in recent decades [11121314], and both receptors are thought to be involved in its adjuvant activity. Flagellin has also been used as a fusion partner to enhance the solubility of many hard-to-express proteins [15]. However, the conjugation effect of flagellin varies depending on the target antigen or the direction of its conjugation [1316]. Therefore, the conjugation of flagellin to different terminals (the N- or C-terminus) of S0 could influence both solubility and immunogenicity.
In our study, we conjugated flagellin from Vibrio vulnificus, FlaB, to the C- or N-terminus of S0 to generate S0-F and F-S0, respectively. We tested the effect of flagellin conjugation to S0 on the enhancement of solubility. To further improve the solubility of each recombinant protein, trigger factor chaperone tig was used, and the solubility of each recombinant protein was assessed in the presence of tig. We also evaluated the ability of the conjugated flagellin to enhance the S0-specific humoral immune response in Balb/c mice and identified different outcomes depending on the direction of fusion. The underlying mechanisms of the consequent results were deduced through in vitro and in vivo experiments. These observations provide crucial information for the development of a novel PED vaccine and flagellin-based immunotherapeutics.
MATERIALS AND METHODS
Animals
Six-week-old female naïve BALB/c mice (17–19 g) were purchased from Samtako (Korea). The mice were maintained under standard pathogen-free conditions on a 12:12 h light/dark cycle (5 mice per cage), monitored three times weekly and provided with free access to food and water during experimentation. All mice were divided randomly into different groups before immunization and adapted for 1 week before any treatments. Animal experiments were performed in accordance with the guidelines for the care and use of laboratory animals under the approval of the animal ethics committee at Seoul National University (No. SNU-161114-4). After the final serum collection of each experiment, the mice were euthanized with CO2 inhalation.
Vector construction
The nucleotide sequence of the spike protein from the PEDV K14JB01 strain (GenBank accession No. KJ623926) was used as a reference to synthesize a partial protein from the S0 (amino acids 25-229) gene. The whole genome sequence from Vibrio vulnificus YJ016 (GenBank accession No. BA000037) was used as a reference to synthesize FlaB. Each sequence was codon-optimized using the table of codon usage in E. coli before synthesis (Genescript, USA; Bioneer, Korea) and PCR-amplified using various primer pairs provided in Table 1.
Table 1
Primer sequences

![]()
The nucleotide sequence of S0 was amplified by PCR using the primers NS0-F/XS0-R and cloned into the vector pET28a(+) (Novagen, USA) using NheI and XhoI restriction enzyme sites (pS0). The FlaB DNA fragment was amplified by PCR using BFlaB-F/XFlaB-R primers and was cloned into pET28a(+). The resulting construct was digested using NheI and BamHI restriction enzymes and ligated with the S0 DNA fragment that was previously PCR-amplified using NS0-F/BS0-R primers (pS0-F). The FlaB DNA fragment, which was PCR-amplified using SFlaB-F/NFlaB-R primers, was cloned into pET28a(+) pretreated with rSAP. The resulting construct, which contained the correct FlaB direction, was digested using NheI restriction enzymes and ligated with the PCR-amplified S0 DNA fragment using NS0-F/XS0-R primers (pF-S0). To express FlaB alone, the FlaB DNA fragment PCR-amplified using NFlaB-F/XFlaB-R primers was cloned into pET28a(+) using NheI and XhoI restriction enzyme sites (pF). As pET28a(+) supports 6His-tag nucleotide sequences on both ends of the multiple cloning site, the resulting recombinant proteins contained an additional 6His-tag on both the C- and N-terminal regions. DNA sequences of the resulting expression vectors were verified by the dideoxy-chain termination method.
Protein expression and purification of recombinant S0, S0-F, F-S0 and F
Each expression plasmid (pS0, pS0-F, pF-S0 and pF) was transformed into E. coli BL21(DE3) (Invitrogen, USA). A seed culture was prepared with a single colony of recombinant E. coli and grown overnight in LB broth containing 50 μg/mL kanamycin. One percent of the seed culture was inoculated into LB broth containing 50 μg/mL kanamycin and grown until the optical density at 600 nm (OD 600) reached 0.5. Cultures were then treated with 0.3 mM IPTG to induce expression of the protein of interest and grown at 25°C for 5 h with shaking set to 200 rpm. Cells were harvested and disrupted on ice by sonication (VCX750, SONICS, USA) using a preset program (45 cycles of 2 sec On/5 sec Off, amp 40%), followed by centrifugation at 20,000 × g for 15 min at 4°C. After centrifugation, supernatants were collected as a soluble fraction, while pellets were solubilized using 50 mM Tris-Cl, 2 M urea, pH 12.5 buffer and collected as an insoluble fraction. Equivalent volumes of solubilization buffer versus supernatants were used to prepare the insoluble fraction. Insoluble and soluble fractions of each recombinant protein were analyzed through SDS-PAGE followed by Coomassie Blue staining (Fig. 1A). The insoluble fraction of S0 was subsequently refolded as described previously [8].
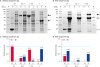 | Fig. 1Analysis of solubility of recombinant proteins in the presence or absence of the chaperone tig. (A, B) Insoluble and soluble fractions of each recombinant protein were analyzed by SDS-PAGE followed by Coomassie Blue staining in the absence or presence of the coexpressed chaperone tig (A and B, respectively). (C, D) Band densities of target recombinant protein from (A) and (B) were analyzed using ImageJ (C and D, respectively). Densities were calculated using the relative ratio of the S0 insoluble fraction without the chaperone tig. Respective values presented for each corresponding bar.IS, insoluble fraction; S, soluble fraction; M, marker; black arrow, location of S0-F or F-S0; dotted arrow, location of F; empty arrow, location of S0; dashed arrow, location of tig.
|
To express each recombinant protein in the presence of the chaperone tig, we transformed each expression plasmid (pS0, pS0-F and pF-S0) into E. coli BL21(DE3)/pTf16 (Takara, Japan), which can induce the expression of trigger factor chaperone tig. A seed culture was prepared by inoculating a single colony in LB broth containing 50 μg/mL kanamycin and 20 μg/mL chloramphenicol overnight at 37°C. tig was expressed before the induction of each recombinant protein. One percent of the seed culture was inoculated into LB broth supplemented with 0.5 mg/ml L-arabinose, 20 μg/mL chloramphenicol and 50 μg/mL kanamycin until the OD 600 reached 0.35. At this point, cells were cooled to 4°C for 15 min and subsequently induced with 0.1 mM IPTG at 15°C for 24 h. The cell disruption process was as described above. Insoluble and soluble fractions of each recombinant protein were analyzed by SDS-PAGE followed by Coomassie Blue staining (Fig. 1B). The band densities of each recombinant protein were measured using Image Lab software (Bio-Rad Laboratories) (Fig. 1C and D).
The soluble fractions of S0-F and F-S0, which were expressed in the presence of tig, were purified using affinity chromatography on a Ni2+-NTA column. Purified samples were dialyzed against phosphate buffered saline (PBS) at 4°C three times and subsequently concentrated using Amicon ultra-15 centrifugal filters (Merck, Germany). Contaminating lipopolysaccharide (LPS) was further removed by using an endotoxin removal spin column (Thermo Scientific, USA), and the residual LPS content of each recombinant protein was determined using the ToxinSensor Chromogenic LAL Endotoxin Assay Kit (Genescript, USA). The LPS levels of each recombinant protein were below 0.03 EU/mL and 0.0075 EU per dose.
IL-8 and MCP-1 release by Caco-2 cells
Caco-2 cells (KCTC, Korea) were seeded at 1 × 105 per well in 48-well plates (SPL, Korea) and incubated at 37°C for 5 days in Dulbecco's modified Eagle's medium (Invitrogen, USA) supplemented with 10% fetal bovine serum (Gibco, USA) in an atmosphere containing 5% CO2. Cells were then cultivated overnight without FBS and subsequently stimulated with different concentrations of each recombinant protein for 6 h. The supernatants were harvested, and cytokine levels were quantified using enzyme-linked immunosorbent assay (ELISA) kits (Cusabio, China).
Serum cytokine detection
Six-week-old BALB/c mice (n = 3) were injected intraperitoneally with 150 pmol of each recombinant protein in 100 μL PBS (3.8 μg of S0, 10 μg of S0-F or F-S0 and 6.2 μg of F per mouse, a total of 4 groups, total n = 12). Sera were collected 1 and 4 h postinjection from intrapetrosal veins with a disposable syringe and delivered into sterilized tubes. Serum was separated by centrifugation at 12,000 rpm for 3 min using a serum separator tube (BD microtainer, USA). Cytokine levels were analyzed using IL-6 and IL-18 ELISA kits (Invitrogen, USA; Cusabio, China, respectively).
Mouse immunization and antibody detection
Six-week-old BALB/c mice (n = 5) were immunized intramuscularly with 30 pmol of each recombinant protein (0.76 μg of S0, 2 μg of S0-F or F-S0 and 0.76+1.24 μg of S0/F per mouse, a total of 5 groups, total n = 25) three times at 2-week intervals after adaptation. CFA and IFA were mixed with antigens for the priming and boosting of mice, respectively. Blood samples were collected on days 13, 27, and 42 (sacrifice) from intrapetrosal veins with a disposable syringe and delivered into sterilized tubes. Serum was separated by centrifugation at 12,000 rpm for 3 min using a serum separator tube (BD microtainer, USA).
To detect S0-specific IgG levels, sera were analyzed by ELISA. 96-well immunoplates (SPL, Korea) were coated with 10 μg/mL of purified recombinant S0 (100 μL/well) in carbonate-bicarbonate buffer (pH 9.6) at 37°C for 2 h. Coated plates were blocked with 0.5% skim milk in PBS (100 μL/well) for 1 h at room temperature. Then, serially diluted serum from a 1:100 dilutions was added to the wells (100 μL/well) and incubated at 37°C for 2 h. After incubation, horseradish peroxidase-conjugated goat anti-mouse IgG, IgG1, IgG2a (1:2,000 dilutions, 100 μL/well) or IgA (1:5,000 dilutions, 100 μL/well) was added to the designated wells for 1 h at room temperature. Appropriate washing of each well was performed between each step with PBS containing 0.05% Tween 20. TMB solution (Sigma-Aldrich, USA) was added to the wells (100 μL/well) as an HRP substrate and incubated for 30 min without light interference, followed by the addition of stop solution (0.16 M H2SO4; 100 μL/well) to stop the enzymatic reaction. Finally, the absorbance was measured at 450 nm using a microplate reader (Infinite® 200 PRO, USA). ELISA results were expressed as the endpoint titer.
RESULTS
Construction, soluble expression and purification of S0-F, F-S0 and F
The truncated S0 region (25-229 amino acid) of the PEDV spike protein, which contains neutralizing epitopes and supports cross-protection among non-S-Indel PEDV strains [10], was chosen as the target antigen. Bacterial flagellin from Vibrio vulnificus, FlaB, was used as a fusion adjuvant candidate. FlaB was fused to the C- or N-terminus of S0 to generate S0-F and F-S0, respectively (Fig. 2A). Nucleotide sequences encoding each recombinant protein (S0, S0-F, F-S0 and F) were inserted into the pET-28a(+) expression vector, which supports hexahistidine tags (6His-tags) that flank the N- and C-terminus of the target protein to enhance purification efficiency. The BamHI and NheI restriction enzyme sites were used to generate S0-F and F-S0 in which each sequence of S0 and F were linked by two amino acids (Gly-Ser or Ser-Ser, respectively). All genetic codons were optimized to facilitate expression in E. coli.
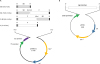 | Fig. 2Schematic view of the expression vector system. (A) Construction of expression vectors for recombinant S0, S0-F, F-S0 and F. (B) Vector map of tig expression plasmid pTF16 purchased from TAKARA BIO INC.S0, truncated region of porcine epidemic diarrhea virus spike protein; F, FlaB from Vibrio vulnificus; H, 6X His-tag.
|
We transformed each expression vector into E. coli BL21(DE3) and induced expression with IPTG. After cell disruption by sonication, soluble lysates were separated from insoluble lysates. Enhancing the solubility of each recombinant protein was deemed an important factor because we planned to utilize the soluble lysates for further experiments. To assess soluble expression, soluble and insoluble fractions of each recombinant protein were confirmed through SDS-PAGE followed by Coomassie Blue staining. S0 was expressed almost entirely as an insoluble protein, while 99% of flagellin was expressed as a soluble protein (Fig. 1A and C). In contrast to F-S0, which was expressed mostly as an insoluble protein, 59% of S0-F was expressed as a soluble protein. Trigger factor chaperone tig was used to enhance solubility, as it has been shown to assist protein folding by protecting long hydrophobic nascent chains during synthesis and initial folding stages [17]. We cotransformed the tig expression vector pTF16 (Fig. 2B) into E. coli BL21(DE3) together with each target protein's expression vector (pS0, pS0-F and pF-S0). Each target protein was induced by adding IPTG in the presence of tig, which had already been induced by L-arabinose addition. In this condition, 66% of S0 was expressed as a soluble protein, in contrast to S0-F and F-S0, which were almost entirely expressed as soluble proteins (Fig. 1B and D). Interestingly, the total amount of F-S0 expression was 6.2 times higher than S0-F. Flagellin appeared to be expressed well in a soluble form without tig assistance. Each target protein in soluble and insoluble fractions was also confirmed by western blot analysis using anti-histag antibody (Supplementary Fig. 1). Data revealed that the soluble expression patterns among target proteins have similar tendency to those of Coomassie blue staining results in Fig. 1.
For further experiments, we purified soluble lysates of each recombinant protein utilizing Ni2+-NTA affinity chromatography, subsequently removed endotoxins and confirmed through SDS-PAGE followed by Coomassie Blue staining (Fig. 3A and C). The representative band of each recombinant protein appeared at the appropriate molecular weight location in the IPTG-induced whole cell lysate lane (Lane 2 of Fig. 3A-C) compared with the IPTG noninduced whole cell lysate lane (lane 1 of Fig. 1A-C). The purities of S0-F, F-S0 and F were 90.3%, 92.6% and 99%, respectively (Table 2). Due to the difficulty of purifying S0 as a soluble lysate, S0 insoluble aggregates were solubilized, refolded and subsequently purified as described previously [8].
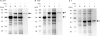 | Fig. 3Purification of S0-F, F-S0 and F. (A) S0-F, (B) F-S0 and (C) F were expressed in E. coli BL21 (DE3). After sonication, soluble fractions were purified. Normalized amounts of each sample were analyzed using SDS-PAGE followed by Coomassie Blue staining.M, marker; W, whole cell lysates; S, soluble fraction; P, purified fraction; black arrows, the location of each recombinant protein; dotted arrow, the location of the chaperone tig.
|

Effect of flagellin conjugation to S0 on the S0-specific humoral immune response
Flagellin's adjuvant effect on the humoral immune response to PED antigen S0 was evaluated through mouse serum S0-specific IgG levels after immunization. Five groups of mice were immunized three times intramuscularly with PBS, 30 pmol of S0 (0.76 μg), S0-F (2 μg), F-S0 (2 μg) and a mixture of S0/F (0.76+1.24 μg) at 14-day intervals. Sera were collected at the 2nd, 4th and 6th weeks postinjection, and S0-specific IgG levels were evaluated by indirect ELISA (Fig. 4A). Interestingly, the F-S0 immunized group had significantly higher levels of S0-specific IgG and IgA compared to all other groups on the 6th week (Fig. 4D and E). Sera collected at the 2nd and 4th week postinjection had a similar tendency as the 6th-week sera (Fig. 4B and C).
 | Fig. 4S0-specific humoral immune response in mice after immunization of each recombinant protein. (A) Schematic view of immunization and blood sampling schedule (n = 5/group) (B-E) Mice were immunized with the indicated recombinant proteins. Blood samples from 2, 4, and 6 weeks postinjection were analyzed with an S0-coated sandwich ELISA. (F) IgG2a and IgG1 subtypes of S0-specific IgG at 6 weeks postinjection. Samples were analyzed by ELISA, and the IgG2a/IgG1 ratio was calculated. All data were plotted on a log 10 scale.ELISA, enzyme-linked immunosorbent assay.
*p < 0.05; **p < 0.01.
|
We also evaluated serum S0-specific IgG2a and IgG1 titers as indicators of Th1 and Th2 type immune responses [18], respectively, at the 6th week postinjection. Data were expressed as a ratio of IgG2a/IgG1. Fig. 4F shows that the S0-immunized group had a more dominant S0-specific IgG1 response than IgG2a response, apart from one mouse. Mice from the S0-F or F-S0 immunized groups also had a more dominant S0-specific IgG1 response than IgG2a response. Interestingly, mice immunized with the S0/F mixture had a more dominant S0-specific IgG2a response than IgG1 response (Fig. 4F).
Evaluation of S0, S0-F, F-S0, F's ability to activate the TLR5 or NLRC4/NAIP5 pathway
Flagellin has been shown to be a highly potent adjuvant through TLR5 in a number of innate and adaptive immune cells [19]. To interpret different S0-specific IgG outcomes among various groups receiving the flagellin adjuvant, we used the epithelial cell line Caco-2, which expresses TLR5 constitutively (Supplementary Fig. 3), to assess the TLR5 activating ability of each recombinant protein. Caco-2 cells were treated with each recombinant protein at 1, 10, or 100 nM. IL-8 and MCP-1, which are representative cytokines for the TLR5 pathway, were measured in the supernatant by ELISA 72 h after treatment. As shown in Fig. 5A, S0-F, F-S0 and F effectively induced IL-8 more significantly than S0 alone at 10 and 100 nM. At 1 nM, treatment with F induced IL-8 more significantly than treatment with S0-F or F-S0. Similar patterns were observed for MCP-1 (Fig. 5B). There was no evident difference in the ability of S0-F and F-S0 to induce IL-8 or MCP-1. For more in-depth analysis, we verified each recombinant protein's ability to activate the TLR5 pathway in vivo. Mice were injected intraperitoneally with equivalent molar values of each recombinant protein, and sera were analyzed by ELISA to detect IL-6 as a marker of TLR5 activation. The data revealed that the F treatment induced stronger serum IL-6 levels when compared to S0-F or F-S0 treatments at 4 h (Fig. 6A). Consistent with the in vitro data, S0-F and F-S0 had a tendency to induce higher levels of serum IL-6 than S0, but the levels were lower than for F. TLR5 stimulation profile of each recombinant protein was also confirmed by TLR5 reporter system and similar tendency was once again confirmed (Supplementary Fig. 2).
 | Fig. 5The TLR5 activation properties of S0, S0-F, F-S0 and F in the Caco-2 model. (A) IL-8 and (B) MCP-1, downstream cytokines of TLR5, were secreted from Caco-2 cells 72 h after treatment with 1, 10, 100 nM of S0, S0-F, F-S0 and F. Supernatants were analyzed by enzyme-linked immunosorbent assay.
*p < 0.05; **p < 0.01; ***p < 0.001, respectively.
|
 | Fig. 6The TLR5 and NLRC4/Naip5 activation properties of S0, S0-F, F-S0 and F in a mouse model. Mice were intraperitoneal injected with equivalent moles of S0, S0-F, F-S0 and F. Blood was collected 1 and 4 h after injection. From blood samples, (A) IL-6 and (B) IL-18, downstream cytokines of TLR5 and Nlrc4/Naip5, respectively, were analyzed by enzyme-linked immunosorbent assay.
*p < 0.05.
|
Flagellin is recognized not only by the transmembrane receptor TLR5 on the cell surface but also by the cytosolic receptor NLRC4/NAIP5 [20]. Flagellin's adjuvant effect on the humoral immune response can be influenced by activation of both receptors [2122]. Therefore, we also sought to determine the ability of each recombinant protein to activate the cytosolic receptor NLRC4/NAIP5 by measuring its responsive cytokine IL-18 from sera. Interestingly, the S0-F-injected group had serum IL-18 levels that were significantly higher than all other groups at 1 h (Fig. 6B). Similar patterns were also observed at 4 h.
DISCUSSION
Flagellin has been frequently used as a molecular adjuvant in conjugation with various antigens [11121314]; however, the adjuvant effect of flagellin varies depending on the target antigen or the position of its conjugation [1316]. In this study, we conjugated flagellin to the N- or C-terminus of PEDV antigen S0 and evaluated its ability to enhance both solubility and the antigen-specific humoral immune response.
Expression of recombinant proteins in an E. coli system has many advantages in terms of cost, yield and scalability; however, expression in E. coli often leads proteins of interest to form biologically inactive aggregates known as inclusion bodies (IBs) [23]. Despite enclosing neutralizing epitopes, PED subunit vaccine candidate antigens, including the S0 (25-229 amino acid), S1D (636-789 amino acid) and COE (499-638 amino acid) regions of the PEDV spike protein, are prone to be expressed as IBs [89]. Although IBs can be refolded by several methods, it is better to express recombinant proteins in a soluble form and prevent IB formation in the first place [232425]. There are various approaches to enhancing the solubility of recombinant proteins, including conjugation of a solubility-enhancing fusion partner protein or optimization of protein expression by adjusting the physiological condition of the host cells [232627]. Here, we tested whether a flagellin fusion could enhance the solubility of the recombinant PEDV antigen S0 in the presence or absence of the trigger factor chaperone tig.
As shown in Fig. 1A and C, and in our previous study [8], the major fraction of S0 is expressed as an IB. In contrast to S0, 59% of S0-F was expressed as a soluble protein, while F-S0 was mainly expressed as an IB. Our data suggest that flagellin conjugation to the C-terminus of S0 improved solubility in the E. coli expression system without tig expression. To improve the solubility of the recombinant proteins, trigger factor chaperone tig, which has been shown to assist protein folding during synthesis and the initial folding process, was introduced and coexpressed with the respective protein of interest. In contrast to S0, which had 66% solubility, 99% of S0-F and F-S0 were expressed as soluble proteins (Fig. 1B and D), suggesting that flagellin conjugation also improved the solubility of recombinant proteins in the presence of tig. Furthermore, coexpression of tig ameliorated the solubility of S0, S0-F and F-S0 from 0% to 66%, 59% to 99%, and 6% to 99%, respectively. Taking this result into account, both flagellin conjugation and chaperone tig coexpression increased the solubility of S0-related recombinant proteins.
Interestingly, despite their similar molecular weights, the total amount of F-S0 expression was approximately 6.2- and 1.5-times higher than S0-F in the presence or absence of tig, respectively (Fig. 1). This difference in protein levels suggests that the N-terminal flagellin sequence may offer efficient translation initiation, resulting in increased production of F-S0. In fact, it is generally accepted that fusion of a solubility enhancer to the N-terminus of a target protein can enable efficient translation initiation, which makes it a more favorable choice for soluble expression than the C-terminal fusion [28]. The conjugation of flagellin to the C-terminus of S0 (S0-F) improved soluble expression in comparison to the N-terminal conjugation (F-S0) or S0 alone in the absence of tig (Fig. 1A and C). tig enhances the solubility of recombinant protein through by assisting with the protein folding process. Proteins that are prone to aggregation, including the S1D and COE region of PEDV's spike protein, which are otherwise totally insoluble, have been expressed as soluble proteins with the assistance of tig [9]. In contrast, the condition lacking tig had a relatively lower capacity to fold recombinant protein correctly and make soluble protein. Therefore, rapid translation of F-S0 in the absence of tig is thought to produce IBs rather than soluble protein in spite of its total protein expression level being 1.5-times higher than the S0-F protein level.
As shown in Fig. 3 and Table 2, S0-F, F-S0 and F underwent purification, resulting in 90.3%, 92.6% and 99% purity, respectively. However, purified S0-F and F-S0 had remarkably lower expression yields than purified F. Differences between the band intensities of the soluble and purified fractions revealed that there was a significant amount of target protein loss during the purification process (lanes 3 and 4 of Fig. 3A-C). We inferred that both 6His-tags bound to the N- and C-termini of S0-F and F-S0 were difficult to access by the Ni2+ metal ions during the purification process.
To protect piglets, the main contributor to substantial economic loss, it is imperative to immunize sows to establish the passive immunity of piglets via colostrum/milk. Therefore, enhancement of the PEDV-specific humoral immune response is a prerequisite for PED vaccine development. Flagellin has been used as a fusion adjuvant for a broad range of recombinant antigens in past decades [111214]; however, there have been relatively few studies assessing the effect of the flagellin fusion position on its adjuvant activity. Therefore, we tested the ability of S0-F and F-S0 to induce S0-specific humoral immune responses in mice. Interestingly, as shown in Fig. 4B-E, we confirmed that flagellin exerts a differential adjuvant effect depending on its conjugation position in reference to S0. The S0-specific antibodies in Fig. 4 is inferred to be the neutralizing antibodies, as the PEDV neutralization titers have positive correlations to indirect ELISA results which use S0 produced from E. coli as a coating protein [29]. We used Complete Freund's adjuvant (CFA) and incomplete Freund's adjuvant (IFA) together with our conjugated vaccine system in all mice immunization experiments equally as a ‘basal’ adjuvant (Fig. 4) because we expected the additional adjuvant effect from the flagellin which could enhance the antigenicity of the S0 over the basal Freund's adjuvant effect.
As flagellin has shown enormous potency as an adjuvant through TLR5 on a number of innate and adaptive immune cells [19], we evaluated the ability of each recombinant protein to activate TLR5 to interpret different S0-specific IgG outcomes among various flagellin adjuvanted groups. As shown in Fig. 5, flagellin, regardless of whether it was conjugated to S0, activated the TLR5 pathway. In particular, F alone activated this pathway to significantly higher levels than the levels achieved by S0-F or F-S0 based on the 1 nM treatment data. This activation tendency was also confirmed in vivo (Fig. 6A). The superior ability of F-S0 to induce S0-specific IgG relative to the mixture of S0/F may result from the effects of the conjugation. In fact, the conjugation of immunopotentiators to antigens has been demonstrated to elicit stronger immune responses than nonconjugated, separated treatments in many previous research investigations [14303132]. This stronger response occurs because conjugation ensures the delivery and uptake of both antigen and immunopotentiator by the same antigen presenting cell, assuring the potentiation of antigen-specific immune responses [333435]. Indeed, the conjugation of flagellin to various antigens has been shown to provide stronger antigen-specific humoral immune responses than separated mixtures [143236].
The cells of the innate immune system are equipped with two germline-encoded receptors to detect flagellin present on flagellated bacteria. The transmembrane receptor TLR5 detects extracellular flagellin, thus activating a number of innate and adaptive immune cells via methods such as secretion of proinflammatory cytokines. Injection of virulence factors containing flagellin from invading microbes results in flagellin being found within the cytosol of macrophages and being recognized by the cytosolic receptor NLRC4/NAIP5, which supports a stronger pathogen killing environment through IL-18, IL-1β and pyroptosis [20]. The role of these two receptors on, in particular, flagellin's adjuvant effect is suggested to be complementary, as a single knockout of either pathway did not interfere with the adjuvancy of flagellin, while a double-knockout resulted in a significant reduction of the adjuvancy of flagellin [2137]. However, the NLRC4/NAIP5 pathway may interfere with flagellin's adjuvancy in the presence of intact TLR5. Li W et al. [22] demonstrated that FliC-L3A flagellin that lacks the ability to activate NLRC4/NAIP5 due to a point mutation, induced a significantly higher antigen-specific serum IgG titer than wild-type flagellin when used as an adjuvant with the model antigen p24 in mice. A/J mice, which are naturally partially deficient in Naip5, displayed higher IgG responses to ovalbumin (OVA) than C57BL/6 mice after immunization with a mixture of OVA and flagellin [37]. Our data revealed that S0-F treatment induced significantly higher levels IL-18, a representative cytokine of the NLRC4/NAIP5 pathway, than F-S0 or F treatment in mice (Fig. 6B), suggesting that the superior ability of S0-F to activate the NLRC4/NAIP5 pathway may interfere with flagellin's adjuvancy and reduce serum S0-specific IgG despite its advantage in the conjugated form.
Flagellin showed a variety of abilities to activate its cognate receptors depending on its conjugation status. In fact, there have been several reports assessing the effect of the directionality of the flagellin fusion on its bioactivity. For example, flagellin conjugated to the C-terminus of HPV-16 E7 protein activated the TLR5 pathway 10-fold more potently than its counterpart, whereas flagellin conjugated to the N-terminus of E7 stimulated the NLRC4/NAIP5 pathway significantly more than its counterpart or even flagellin alone [16]. We speculate that the receptor binding domains of flagellin may be influenced by the presence of the S0 protein, causing the NLRC4/NAIP5 binding domain of S0-F to be more exposed than the binding domain F-S0 or F.
Here, we evaluated the adjuvant effect of flagellin by the induction of systemic humoral immune response (Fig. 4). Though many studies immunized the sow through intramuscular route [38], induction of the virus-specific IgA at colostrum/milk resulted from mucosal immunization is important factor in terms of lactogenic immunity [39]. Further works using target animals, however, is crucial for the development of this antigen as an effective vaccine.
In this study, we conjugated flagellin, Vibrio vulnificus FlaB, to the N- or C-terminus of PEDV antigen S0 and evaluated its ability to improve protein solubility and an antigen-specific humoral immune response. The ability of conjugating an immunopotentiator to an antigen to enhance the performance of the vaccine was once again confirmed. Conjugation of flagellin to S0 improved the solubility of the recombinant proteins when combined with chaperone tig. Flagellin conjugation did not improve the solubility of all other proteins; however, flagellin has been shown to be a good candidate for enhancing antigen solubility in proteins that are prone to form IBs such as S0. Conjugation of flagellin to the N-terminus of S0 enhanced the S0-specific humoral immune response more than any other type of flagellin assistance. However, flagellin conjugation to the N-terminus of other antigens (e.g. HIV gp120, HPV E7) is not guaranteed to improve the antigen-specific humoral response because flagellin's preference to activate TLR5 or NLRC4/NAIP5 in its position differs depending on the conjugated antigens. Rather, we suggest that flagellin-conjugated antigens, which have an inferior ability to activate NLRC4/NAIP5, may be good candidates for inducing an increased antigen-specific humoral immune response. Furthermore, S0-induced responses were dominated by S0-specific IgG1 rather than IgG2a. Because the induction of IgG2a and IgG1 represent Th1- and Th2-type immune responses, respectively [18]. S0 seems to have an inherent ability to induce Th2-type immune response to itself (Fig. 4F). In contrast to S0-F and F-S0, flagellin alone induced IgG2a more than IgG1, suggesting that flagellin skewed the type of immune response from Th2 to Th1. As Th2 cell-derived cytokines, such as IL-5 and IL-6, are important for the generation of IgA at mucosal site [40], S0-F or F-S0 is expected to have an additional advantage if they are utilized as a mucosal vaccine. These findings provide crucial information for the rational design of a PED vaccine and flagellin-based immunotherapeutics.
 XML Download
XML Download