

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Schmallenberg virus (SBV), a member of the Peribunyaviridae family, genus Orthobunyavirus [1], is an arthropod-borne pathogen that is transmitted by biting midges (Culicoides spp.) [2]. Discovered in Germany in 2011 [34], this virus has since spread throughout the European continent [5]. The disease primarily affects ruminants, including cattle, sheep, and goats. A SBV infection is associated with diarrhea, fever, and decreased milk production [3678], as well as widespread abortions and developmental malformations in newborn domestic ruminants [9]. Inactivated vaccines have been developed to help prevent the economic losses associated with SBV infections and are commercially available [101112], but these vaccines do not offer the possibility of differentiating infected from vaccinated animals (DIVA).
The SBV genome, similar to other orthobunyaviruses, has a tripartite single-stranded RNA genome of negative polarity consisting of small (S), medium (M), and large (L) RNA segments. The L segment encodes the RNA-dependent RNA polymerase [1314], and the S segment encodes the nucleocapsid (N) protein and nonstructural protein NSs. Commercial N-based enzyme-linked immunosorbent assay (ELISA) tests are available for SBV antibody detection [1516] and they are used extensively for surveillance. The M segment encodes 2 structural glycoproteins, amino-terminus glycoprotein (Gn) and carboxyl-terminus glycoprotein (Gc), as well as the nonstructural protein, NSm. The 2 orthobunyavirus glycoproteins differ in size; Gn is 32–35 kDa, whereas Gc is 100–110 kDa. Both proteins are type I integral membrane proteins modified by N-linked glycosylation, and are rich in cysteine residues. The humoral immune response against mammalian cell-expressed SBV glycoproteins has been analyzed and only Gc, not the Gn protein, was recognized by SBV hyperimmune sera in various serological assays [17].
The authors' previously reported a recombinant subunit vaccine for Rift Valley fever (RVF) using baculovirus expressed RVF virus (RVFV) Gn and Gc glycoproteins that resulted in excellent protection from a virulent RVFV challenge [18]. The amino acid (aa) domains associated with SBV neutralization have recently been identified to be located within the SBV Gc protein. Previously, different SBV subunit vaccines, containing the full ectodomains of both Gc and Gn, or the critical aa domain of Gc expressed in mammalian cells or as DNA plasmid vaccine, were used in cattle and IFNAR−/− mice, resulting in partial protection; not all vaccinated cattle were protected from developing viremia and not all mice were protected from mortality [1920]. Using a similar strategy to that with the RVF subunit vaccine, the ectodomains of SBV Gc and Gn were expressed in baculovirus and these proteins were used as candidate vaccines to immunize cattle in a prime-boost approach. The immunogenicity and induction of neutralizing antibodies, and subsequent prevention of viremia and tissue infection after SBV challenge were assessed.
MATERIALS AND METHODS
Construction of recombinant baculoviruses expressing SBV Gn and Gc
The cloning and construction of the recombinant baculoviruses was carried out in a similar manner to that described previously [18]. Briefly, the coding sequences of the SBV structural proteins Gn and Gc were obtained from the SBV BH80/11-4 isolate (GenBank accession No. HE649913.1), and synthesized (GENEWIZ, USA) with molecular modifications. For both proteins, the transmembrane domain and cytoplasmic tail were deleted from the coding sequence and a C-terminal 6xHistag was added to facilitate protein purification by affinity chromatography using the Ni-NTA superflow resin (Novagen, USA).
Expression and purification of SBV Gn and Gc proteins
Purified recombinant bacmids carrying the respective coding sequences of SBV Gn and Gc were transfected into Sf9 cells (Invitrogen–Life Technologies, USA) using the Cellfectin II Reagent (Invitrogen–Life Technologies), grown in Sf-900 II SFM medium (Invitrogen–Life Technologies), and supplemented with 10% fetal bovine serum and 100 U/mL penicillin-streptomycin. Protein expression was carried out using a P4 passage recombinant baculovirus stock (> 107 plaque-forming units [pfu]/mL). Western blot analysis of the lysate was performed to confirm the SBV-specific protein expression, as described below. The recombinant SBV proteins were purified using Ni-NTA Super flow resin according to a protocol described previously [21]. The protein concentration was determined using the bicinchoninic acid assay (Thermo Fisher Scientific, USA) at an absorbance of 562 nm using bovine serum albumin (BSA; Sigma-Aldrich, USA) as the protein standard. Aliquots were stored at –80°C until needed.
Western blot analysis to detect recombinant SBV Gc and Gn proteins
The detailed procedure for detection of baculovirus-expressed recombinant proteins by western blot is described elsewhere [21]. Briefly, the proteins were separated using a 12% Bis-Tris gel (Life Technologies) and transferred to a polyvinylidene fluoride membrane (Millipore, USA). The membrane was blocked with 0.05% Tween-20 in 1× PBS (pH 7.4) containing 3% BSA for 1 h, and incubated overnight at 4°C with mouse anti-histidine horseradish peroxidase-conjugated monoclonal antibodies at a dilution of 1:5,000 (Thermo Fisher Scientific). After a final wash steps with 1× PBS, the specific reactivity was detected using a 3-amino-9-ethyl-carbazole peroxidase substrate (Sigma-Aldrich) detection system.
Vaccine preparation
The purified SBV Gn and/or Gc glycoproteins were formulated according to the manufacturer's instructions in Montanide ISA 25 oil-in-water adjuvant (Seppic, France) to obtain a concentration of 100 or 150 µg of each protein per vaccine dose in the first and second vaccine trials, respectively. The vaccines were administered subcutaneously.
Animals and experimental design of challenge studies
The experiments involving animals and infectious SBV were carried out at the Biosecurity Research Institute of Kansas State University in accordance with the animal welfare guidelines and were approved by the Kansas State University's Institutional Animal Care and Use Committee (IACUC No. 3575). Cattle were selected as the target animal for the vaccine trials based on previous studies revealing more obvious virological and serological responses in cattle than sheep [22]. Twenty-four, 6-month-old, Holstein-Friesian cattle were purchased locally from a commercial farm and used in 2 separate studies. In both studies, the animals were assigned randomly to the different experimental groups using a random number table. The first vaccine trial had 2 groups (n = 6 each) of cattle. Group 1 was administered Gc (100 µg/dose) and group 2 was administered PBS, subcutaneously. The animals were boosted with the same dose 2 weeks after the first vaccination and challenged 9 days after the booster vaccination with 1 mL of bovine serum containing 104 pfu SBV strain BH80/11-4, subcutaneously (Fig. 1A).
 | Fig. 1Experimental design. (A) Trial 1, 12 calves were assigned to 2 groups (G1-1 received 100 µg of SBV Gc, G1-2 received PBS); (B) Trial 2, 12 calves assigned to 3 groups (G2-1 received 150 µg of SBV Gc, G2-2 received SBV Gc/Gn (150 µg each), and G2-3 received PBS. Booster injections were given after the 2nd and 3rd week post-first vaccination in trials 1 and 2, respectively. Vaccines and PBS were prepared with adjuvant.SBV, Schmallenberg virus; dpv, days post-vaccination; Gc, carboxyl-terminus glycoprotein; Gn, amino-terminus glycoprotein; PBS, phosphate buffered saline.
|
The second vaccine trial had 3 groups (n = 4 each) of cattle: group 1, administered Gc (150 µg/dose), group 2 administered Gc and Gn combined (150 µg of each/dose), and group 3 administered PBS, all subcutaneously. A booster injection of the same amounts of the respective antigens was given 3 weeks following the first vaccination. The unvaccinated control group received 1 mL of PBS mixed with the adjuvant. The cattle were challenged as described for the first trial 3 weeks after the second vaccination (Fig. 1B). At the end of the experiments (necropsy on day 7 for trial 1 and day 8 for trial 2), all animals were euthanized humanely by an intravenous administration of Fatal-Plus (Vortech Pharmaceuticals) at a dose of 1 mL per pound of body weight.
Blood specimen collection
Blood specimens were collected from the jugular vein of all animals prior to vaccination (day 0) to establish a baseline pre-vaccination immune response status. Thereafter, serum samples were prepared from blood of all cattle on days 7, 14, 23, and 30 (first trial), and 14, 21, 28, 35, 42, and 49 (second trial) post-vaccination (dpv). The sera were stored at –20°C until needed. The SBV positive control serum (21 days post-infection cattle serum) was obtained from a previous study [22].
ELISA analysis
The specific antibody response in serum was measured using an indirect ELISA based on the baculovirus-expressed SBV Gc protein. The ELISA assay was performed using a previously described methodology [1518]. The cutoff point was determined by the addition of 2 standard deviations to the corresponding mean OD value of the pre-vaccination serum. The mean OD values equal to or greater than the cutoff value were considered positive.
SBV neutralizing antibodies
SBV neutralizing antibody titers in the serum samples were measured using a plaque reduction neutralization test (PRNT) as described previously [23] with minor modification. Briefly, 2.4% microcrystalline cellulose Avicel RC (FMC BioPolymer, USA), mixed with an equal volume of 2× minimum essential media (MEM) containing 10% fetal calf serum, was used as the overlay. The SBV stock virus (106 pfu/mL) grown in BHK21 cells, was diluted to 50 pfu/well for use in the PRNT assay. Equal volumes of test serum and virus (125 µL each) per well in a 96-well plate were incubated at 37°C for 1 h before being transferred to 80% confluent Vero-E6 cells, and grown in a 24-well plate. Ten 2-fold dilutions were tested in duplicates for each sample. Negative and positive sera, as well as cell control wells were included. The neutralization titer is expressed as the reciprocal of the highest serum dilution yielding ≥ 80% reduction in the number of plaques. The results are presented as the average titer (log2) of each group at different time points post infection.
RNA extraction
Tissue samples (spleen, cervical, mandibular, mesenteric lymph nodes, and Peyer's patches) collected at necropsy were homogenized in 1 mL MEM per 100 mg of tissue using stainless steel beads in the Tissuelyser II (Qiagen, USA). A 250 µL aliquot of tissue lysate was added to 750 µL of Trizol LS reagent and incubated for 5 min at room temperature. The collected serum was treated similarly. Bromochlorophenol (100 µL) was added to the mixture to separate the aqueous phase. The samples were shaken vigorously for 15 sec and incubated at RT for 5 min. The mixture was centrifuged at 12,000 RCF for 15 min at 4°C and the aqueous layer was transferred to a clean tube. For RNA extraction, 100 µL of the aqueous layer was used for both of the following 2 extraction protocols. A King Fisher automated magnetic particle processor (Thermo Fisher Scientific) and the MagMAX Viral RNA extraction kit (Ambion, USA) in trial 1, and a Qiagen BioSprint 96 automated magnetic particle processor (Thermo Fisher Scientific) and the MagAttract 96 cador pathogen kit (Qiagen) in trial 2 were used according to the manufacturer's recommendations.
Reverse transcriptase real time polymerase chain reaction (RT-qPCR)
The SBV genome was detected by S segment-specific real-time reverse transcription quantitative PCR (RT-qPCR), as described previously [24] with the following modifications. In trial 1, the RT-qPCR assay was performed using an AgPath-ID One-Step RT-PCR kit (Applied Biosystems, USA) with a total reaction volume of 25 μL. In trial 2, amplification was carried out with 20 µl using qScrip XLT One-Step RT-qPCR Tough Mix master mix (Quanta Biosciences, USA). To each 20/25 µL reaction volume, 5 µL of RNA template was added. The thermocycling conditions were 50°C for 20 min, 95°C for 5 min, followed by 45 cycles of 95°C for 10 sec and 60°C for 1 min. Each sample was tested in triplicate and both positive and negative controls were included in each run. The cut-off cycle threshold (Ct) value was set to 34 based on the Ct value of the pre-SBV inoculation serum.
RESULTS
Expression of recombinant SBV Gc and Gn proteins
Expression of the SBV Gc and Gn proteins was confirmed by western blot assay using mouse monoclonal antibodies directed against the C-terminal histidine tags of the recombinant proteins. The SBV Gc and Gn proteins showed specific bands in the western blots that matched the expected molecular weight of the recombinant proteins (Fig. 2).
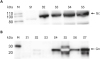 | Fig. 2SBV glycoprotein expression. Recombinant SBV Gc and Gn proteins expression was confirmed by western blot assay using anti-histidine monoclonal antibodies; S1–S7 indicating different batches of protein preparations (A) Gc, (B) Gn, lane S5 appears unclear because of the amount of protein present and high saturation of the image.M, molecular weight marker; S, different batches of protein specimen; Gc, carboxyl-terminus glycoprotein; Gn, amino-terminus glycoprotein; SBV, Schmallenberg virus.
|
Clinical observations after vaccination and challenge
None of the animals in trials 1 or 2 had any adverse reaction to vaccination except for mild localized swelling detected at the vaccination site, which resolved quickly and was most likely associated with the vaccine/adjuvant depot. No clinical signs attributable to the SBV infection were observed after challenge, not even in the unvaccinated challenge control. Most of the cattle (9 out of 12) in trial 1 had clinical coccidiosis and were treated with sulfaquinoxaline (6 mg/lb/day for 5 days). The treatment was successful and symptoms, such as diarrhea and loss of appetite, disappeared within several days of treatment during the vaccination phase of the study.
Serology
Serum was collected weekly and analyzed for SBV-specific antibodies using an indirect ELISA against the SBV Gc protein, the common component of both vaccines, which was used to compare the antibody responses between the 2 different vaccine groups. The SBV Gc-specific antibody response was first detected in the serum samples by 14 dpv for all vaccinated groups. In the first trial, the antibody level increased until the day of virus challenge on 23 dpv (Fig. 3A). In the second trial, the antibody response significantly increased in both the Gc and Gc/Gn groups from days 14 to 28 and then decreased by 42 dpv (Fig. 3B). No antibody response was detected in the mock-vaccinated control groups before the challenge infection on 23 (first trial) and 42 (second trial) dpv. An increase in the Gc-specific antibody reactivity was observed in all animals 1 week after SBV challenge (Fig. 3).
 | Fig. 3SBV-specific antibody response. Results of serum samples tested by Gc-specific indirect enzyme-linked immunosorbent assay in trial 1 (A) and 2 (B). The plates were coated with the recombinant SBV Gc protein. The cutoff point (A = 0.8, B = 0.6) was determined by the addition of 2 standard deviations to the corresponding mean OD value of the pre-vaccination serum.Gc, carboxyl-terminus glycoprotein; Gn, amino-terminus glycoprotein; dpv, days post-vaccination; SBV, Schmallenberg virus.
|
To measure the vaccine-induced neutralizing antibody response, PRNT80 was performed with all sera in 3 replicates for each time point collected, and the average titers are presented in Fig. 4. SBV positive and negative sera were used as controls. In trial 1, a low PRNT80 titer of 1:4 of SBV-specific neutralizing antibodies was detected in 3 out of 6 animals in the Gc-vaccinated group at 14 dpv. The titers increased in 4 out of 6 animals to 1:8 (2/6) and 1:16 (2/6) by the day of challenge at 23 dpv. Fig. 4A shows the average titer for each group in trial 1 for the different time points post infection. Similarly, in the second trial, a titer of 1:4 was present in 14 dpv sera in 2 out of 4 animals in both the Gc- and Gc/Gn-vaccinated groups. By the day of challenge at 42 dpv, titers of 1:8, 1:8, 1:16, and 1:16 for the Gc-vaccinated group (n=4), and 1:4, 1:8, 1:16, and 1:16 for the Gc/Gn-vaccinated group (n=4) were observed. Fig. 4B shows the average titer for each group in trial 2. Neutralizing antibodies were detected in all animals, including the mock-vaccinated controls at 7 days after the challenge infection.
 | Fig. 4SBV neutralizing antibody response. Antibody titers in cattle following vaccination and challenge measured by PRNT80. Average titer (log2) of each group in trials 1 (A) and 2 (B) at different time points post-vaccination.Gc, carboxyl-terminus glycoprotein; Gn, amino-terminus glycoprotein; PRNT, plaque reduction neutralization test; dpv, days post-vaccination.
|
Viremia/RNAemia
Because a SBV infection in non-pregnant animals is mostly subclinical, the detection of SBV RNA in the sera of infected animals is a critical parameter for evaluating the vaccine efficacy after a challenge infection [111920]. Therefore, RT-qPCR was performed on the serum samples from 0 to 7 days post-challenge (dpc) and on tissue samples obtained at necropsy. SBV RNA was detected in the serum from animals in all groups from trials 1 and 2, starting from 1 or 2 dpc with a peak at 3 and/or 4 dpc. SBV RNA remained detectable until 4 dpc, independent of the vaccination status, and became negative at 5 dpc (Fig. 5A and C). In addition, SBV RNA was detected in all tissues collected: cervical, mandibular, mesenteric lymph nodes, spleen, and Peyer's patches. The mesenteric lymph node had the highest SBV RNA load (lowest Ct value) in both trials compared to the other tissues (Fig. 5B and D).
 | Fig. 5Results of the S segment based real-time reverse transcription quantitative polymerase chain reaction. SBV RNA detection in serum and different tissue samples from SBV-infected cattle, vaccinated with SBV Gc, SBV Gc/Gn, or mock-vaccinated for trial 1 (A, B) and 2 (C, D) (Ct cutoff value = 34). Tissues (B, D) were collected at time of necropsy at 7 dpc for trial 1 (B) and 8 dpc for trial 2 (D).SBV, Schmallenberg virus; Gc, carboxyl-terminus glycoprotein; Gn, amino-terminus glycoprotein; dpc, days post-challenge; Ct, cycle threshold.
|
DISCUSSION
SBV vaccines based on inactivated virus formulations have been developed and used to effectively prevent RNAemia, clinical disease, and fetal malformations and premature births or stillbirths [12]. Although inactivated vaccines are considered safe under field conditions, they usually induce weaker or shorter-lived immunity than live vaccines. Modified live virus (MLV) vaccines induce strong immune responses but have safety concerns, which include the risk of contamination with virulent strains or other unwanted pathogens as well as the potential for reversion to virulence [2526]. Furthermore, at the time of this study, none of the SBV vaccines available were DIVA compatible. More recently, an efficacy study using a recombinant MLV vaccine with a NSs deletion in the S segment, capable of DIVA, revealed good protection from SBV infections [11]. However, the safety of this MLV vaccine is of concern because its serial passages in cell culture restored virus virulence, based on a single substitution acquired in the Gc protein [27].
In regards to immune responses against SBV-infections, it is not clear yet what role individual SBV-proteins and the cellular and humoral immune response play in protective immunity and virus clearance [19]. Our study focused on the humoral component of the immune response reported to play a major role in protection. The development of neutralizing antibodies that block SBV infections or viremia, which is associated with protection [28], is a key attribute for vaccines against Simbu serogroup viruses, including SBV [12]. Animals vaccinated with both the Gc and Gc/Gn subunit vaccines seroconverted by 14 dpv and exhibited a time-dependent increase in antibody reactivity in both trials. This is in agreement with other reports, where vaccine preparations containing a peptide subdomain of Gc induced time-dependent SBV-specific antibody responses [1920]. Low PRNT80 titers of SBV-specific neutralizing antibodies were detected in 4 out of 6 animals (titers 1:8 to 1:16) in the Gc group of trial 1, and in 8 out of 8 Gc- or Gc/Gn-vaccinated animals (titers 1:4 to 1:16) in trial 2 at the day of challenge. Surprisingly, SBV RNA was detected in the serum and tissues of these SBV-antibody positive animals by RT-qPCR, indicating an absence of protection against a SBV infection. No significant difference in RNAemia levels between the vaccinated and mock-vaccinated animals was noted. This might be because the titers of neutralizing antibodies were too low and not sufficient to protect animals from SBV challenge infection. A similar vaccination strategy was used in a study which reported that despite neutralizing antibodies present after vaccination with the full ectodomains of the Crimean-Congo hemorrhagic fever virus Gn and Gc glycoproteins (also expressed in insect cells), protection against challenge with this bunyavirus was not achieved [29]. Interfering effects of the clinical coccidiosis observed during the vaccination phase of the first trial was ruled out due to similar results obtained in the second trial, where the animals were free of this parasite.
Neither a DNA vaccine encoding the SBV Gc amino-terminal domain (aa 678-947) nor a vaccine based on the Escherichia coli expressed recombinant SBV Gc amino-terminal domain (aa 468-702) conferred very good protection against SBV infection in a mouse model [1920]. Using the DNA vaccine, prevention of weight loss and reduced viremia relative to the unvaccinated controls were reported [19]. Interestingly, partial protection was observed when the SBV Gc amino-terminal domain (aa 468-702) was expressed in human embryonic kidney cells and used as a vaccine formulation [20]. Subunit vaccines containing the full ectodomains of covalently linked Gc and Gn or subdomains of Gc have been shown to confer only partial protection (1 out of 4 animals protected); however, full protection was obtained when the amino-terminal domain of SBV Gc was linked to the corresponding domain of the related Akabane virus [20]. A recent report indicated that the SBV Gc amino-terminal domain delivered by recombinant Equine Herpes Virus 1 and Modified Vaccinia Virus Ankara conferred partial or full protection, respectively [30]. These variations observed in the responses against SBV subunit vaccines could be explained by the different antigen designs and preparations which might modify the accessibility of the protective epitopes of the Gc protein used in each vaccine preparation (full ectodomain vs. amino-terminal domain). Furthermore, differences in expression systems (mammalian cells vs. baculovirus vs. E. coli vs. virus vector) can affect post-translational modifications and folding of the proteins, which in turn affect the immunogenicity of the antigen and the specificity of the antibody response elicited by the antigen [31]. The low neutralization antibody titers and lack of protection in the current studies using SBV Gc and Gc/Gn subunit vaccines contrast with the previously reported high efficacy of a Gn and Gc-based subunit vaccine for another bunyavirus, RVFV [32]. The conclusion that SBV Gn has no additional effect on inducing SBV-specific antibodies when used in the combined Gc/Gn vaccine formulation cannot be made because the generation of Gn-specific antibody responses was not tested. On the other hand, analysis of the purified Gn protein by an immunoblot similar to the Gc protein (Fig. 2), suggested efficient expression of the recombinant Gn with the expected molecular weight. Overall, based on these results, neither Gc alone nor a combination of Gc and Gn induced sufficient neutralizing antibody titers to offer protection against a SBV challenge infection in cattle. More studies will be needed to determine the correlates of protection to develop effective vaccines for SBV.
 XML Download
XML Download