

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Renal ischemia and reperfusion (I/R) injury is a common problem in clinical surgeries, such as renal transplantation, and prolonged ischemia results in irreversible damage to renal function.1 Studies have reported that apoptosis is essential to the pathogenesis of renal injury secondary to I/R.23 Proximal tubular epithelial cells (PTCs) are particularly vulnerable to hypoxia-induced injury. More importantly, the vital pathogenesis of renal I/R has been widely considered as renal tubular epithelial cells apoptosis.4 The close connection between renal tubular epithelial cell apoptosis and renal I/R injury has drawn substantial attention in developing anti-apoptosis therapies.
Propofol (2,6-disopropylphenol), an intravenous sedativehypnotic agent well-known for its utilization in anesthesia, has also been demonstrated to alleviate I/R injury due to its anti-inflammatory and anti-oxidant activities.5 However, the precise mechanism thereof remains largely unresolved. Meanwhile, autophagy is an evolutionarily conserved multi-step process that maintains normal function and the structure of the cell and is reported to be involved in renal I/R injury.6 Recent research reported that propofol attenuated hypoxia/reoxygenation-triggered neuronal injury via inhibition of autophagy.7 However, little is known about the underlying mechanisms by which propofol regulates autophagy.
c-Jun N-terminal kinase (JNK, including JNK 1, 2, 3), also known as stress-activated protein kinase, is another subclass of the mitogen-activated protein kinase signaling pathway in mammalian cells. JNK is closely related to inflammation, cell apoptosis, and autophagy,8 and excessive activation of JNK has been implicated in nephropathy and renal I/R injury.910 Accordingly, we hypothesized that propofol could attenuate hypoxia/reoxygenation-induced apoptosis and autophagy in human renal proximal tubular cells (HK-2) by inhibiting JNK activation.
MATERIALS AND METHODS
Cell culture and treatments
HK-2 cells were purchased from Procell Life Science & Technology Co., Ltd. (Wuhan, China). The cells were cultured in minimum essential medium supplemented with 10% fetal bovine serum both from Gibco/Thermo Fisher Scientific (Shanghai, China) and 1% penicillin and streptomycin under a humidified atmosphere consisting of 5% CO2 and 95% air at 37℃ (control group). Hypoxia and reoxygenation (H/R) injury was introduced by exposing the cells to hypoxic conditions (1% O2, 5% CO2 and 94% N2) for 15 h, followed by reoxygenation under normoxic conditions (reoxygenation) for 2 h in fresh normal medium (H/R group).
Propofol (Aladdin Reagent Int., Shanghai, China) was dissolved in dimethyl sulfoxide (DMSO) and subsequently diluted in minimum essential medium to final concentrations of 10, 25, 50, and 100 µM prior to H/R injury (propofol groups); the final concentrations of DMSO were below 0.01%, thereby minimizing its effects. A positive control was set up using cells pretreated with 10 µM SP600125 (MedChemExpress, Shanghai, China), which is a specific JNK inhibitor. Finally, a combination of 50 µM propofol and 10 µM SP600125 was added to the cell cultures at 1 h before H/R injury to ascertain synergistic effects.
Cell viability analysis
Cell viability was assessed with Cell Counting Kit-8 from Biosharp (Guangzhou, China). Briefly, after reoxygenation for 2 h, each well was supplemented with 10 µL of CCK-8 solution, followed by 4 h of incubation at 37℃. Subsequently, optical density of each well was measured with a microplate reader at a wavelength of 450 nm (MULTISKAN MK3, Thermo, USA). Higher cell viability leads to higher optical density reading.
Measurement of lactate dehydrogenase by ELISA
A lactate dehydrogenase (LDH) release assay (Changchun Huili Biotech Co., Ltd, Changchun, China) was used to determine the extent of cell injury by measuring the amount of LDH released into the medium by the cells following the manufacturer's guidance.
Flow cytometry analysis
Flow cytometry analysis was carried out to assess apoptotic index values in HK-2 cells. Following treatment, HK-2 cells were detached from wells by addition of 0.25% trypsin, resuspended in phosphate-buffered saline (PBS), and spun down at 1500 rpm for 5 mins. Subsequently, the cells were resuspended in 500 µL of binding buffer, followed by incubation with Annexin V-FITC (5 µL) and propidium iodide (PI, 5 µL) for 10–15 mins at room temperature (20–25℃) in the dark. Apoptotic cell ratio was detected using a flow cytometer (CytoFLEX, Beckman Coulter Life Sciences, Indianapolis, IN, USA). Early apoptotic cells were differentiated by unique characteristics: positive for Annexin V-FITC and negative for PI.
Western blotting
Cell cultures were harvested and homogenized in ice-cold radioimmunoprecipitation assay buffer with protease inhibitors (Beyotime, Shanghai, China) and incubated for 30 mins at 4℃. After centrifugation, the supernatants, which contained our target proteins, were transferred to a new vial for storage at −70℃ or immediate use. A BCA protein assay kit was then used to quantify the concentrations of proteins (Beyotime, Shanghai, China). The proteins (40 µg) were run on 12% SDS-PAGE gel and then transferred electrophoretically to a polyvinylidene fluoride membrane (Millipore, Shanghai, China). The blots were blocked for 2 h at 25℃ with 5% skim milk in Tris-buffered saline containing 0.1% Tween 20 (TBST), followed by incubation with primary antibodies, including anti-JNK, anti-p-JNK, anti-caspase-3, anti-Bcl-2, anti-LC3, anti-p62, and anti-β-actin antibodies (Proteintech Group, Inc., Wuhan, China) at 4℃ overnight. Subsequently, membranes were washed with TBST five times for 5 mins each. Then, respective horseradish peroxidase-linked secondary antibodies (Beyotime, Shanghai, China) were added and incubated for 2 h at 37℃, followed by washing with TBST. Lastly, ECL solution was applied to the membrane evenly prior to detection of chemiluminescence (Thermo Scientific, Shanghai, China).
Immunofluorescence assay
The HK-2 cells were fixed in 4% paraformaldehyde for 15 mins and washed three times with PBS for 3 mins each time. Then, the samples were treated with 0.5% Triton X-100 for 20 mins, washed, and blocked for 30 mins at 37℃. Next, the samples were incubated in a wet box at 4℃ overnight with primary antibodies against LC3 (mouse anti-LC3 antibody or p62 (rabbit anti-p62 antibody) diluted 100 times in blocking solution. After washing with PBS containing 0.1% Tween 20, the cells were incubated with secondary antibodies diluted 100 times (LC3, Cy3-labeled goat anti-mouse IgG; p62, Cy3-labeled goat anti-rabbit IgG) for 1 h in the dark at 37℃. Lastly, 4,6-diamidino-2-phenylindole (Beyotime, C1002) was used as a counterstain. After staining, cells were visualized with a fluorescence microscope (BX53, Olympus, Tokyo, Japan).
Statistics analysis
Statistical analyses were conducted with GraphPad Prism version 6 (GraphPad Software Inc., San Diego, CA, USA). Each experiment was repeated six times. Results are presented as mean±standard deviation (SD). Statistical significance was analyzed first with one-way analysis of variance and then with Tukey's multiple comparisons test. p values less than 0.05 were considered to represent statistically significant differences.
RESULTS
Propofol alleviates the reduced cell viability induced by H/R injury
In the present study, we investigated the effect of H/R insult on HK-2 cells. As shown in Fig. 1, H/R decreased cell viability by 39%, compared with the no-insult control group (p<0.001). Propofol significantly alleviated the decrease in cell viability induced by H/R insult in a dose-dependent manner (H/R vs. Pro 10, p=0.045; H/R vs. Pro 25, p<0.001; H/R vs. Pro 50, p<0.001; H/R vs. Pro 100, p<0.001). However, a peak increase in cell viability was observed with pretreatment of propofol at a dose of 50 µM, probably due to the saturation of ligand binding (Fig. 1).
Propofol attenuates LDH release induced by H/R injury in HK-2 cells
We then evaluated the effects of propofol on LDH release in HK-2 cells. As shown in Fig. 2, H/R increased LDH release 3.8-fold over that in the control group, which was significantly attenuated by propofol in a dose-dependent manner. Maximal attenuation of LDH release (1.5-fold that of the control group) was observed at a dose of 50 µM propofol. Interestingly, 100 µM propofol seemed to increase LDH leakage, compared to 50 µM propofol, although there was no statistical difference (p>0.05). The possible cause was related to cytotoxicity from the high concentration of propofol (Fig. 2).
Propofol pretreatment attenuates H/R induced cell apoptosis in HK-2 cells
The effects of propofol pretreatment were also determined by detecting cell apoptosis using flow cytometry. As shown in Fig. 3, H/R injury lead to a 3.5-fold increase in cell apoptosis, compared to the control group (p<0.001), and this alteration was reduced by propofol treatment (H/R vs. Pro 10, p=0.038; H/R vs. Pro 25, p=0.003; H/R vs. Pro 50, p<0.001; H/R vs. Pro 100, p<0.001). Consistently, peak reduction of cell apoptosis was obtained with treatment of 50 µM propofol. Based on the above results, we chose 50 µM propofol for subsequent experiments.
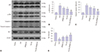
Propofol pretreatment attenuates H/R-triggered cell apoptosis of HK-2 cells by inhibiting JNK activation
Given the essential role of JNK signal transduction pathway in cell apoptosis, we hypothesized that propofol would alleviate H/R-induced cell apoptosis in HK-2 cells by inhibiting JNK activation. To test this, we first ascertained the percentages of apoptotic cells in total cell cultures by measuring Annexin V-FITC and PI with flow cytometry (Fig. 4). As expected, we found that H/R injury induced cell apoptosis, while pretreatment with 50 µM propofol alleviated the induced cell apoptosis (p<0.01). Similar effects were also observed with pretreatment of 10 µM SP600125 (p<0.01). Interestingly, a combination of 50 µM propofol and 10 µM SP600125 offered stronger protection against cell apoptosis (p<0.01), indicating potentially synergistic effects (combination of 50 µM propofol and 10 µM SP600125 versus 50 µM propofol, p<0.05) We also examined the protein expressions levels of JNK, p-JNK, cleaved caspase-3, and Bcl-2 by Western blotting. As shown in Fig. 5, H/R injury resulted in elevated protein levels of p-JNK and cleaved caspase-3 and decreased protein levels of Bcl-2 (all p<0.01). All these changes were attenuated by pretreatment with 50 µM propofol or 10 µM SP600125 (all p<0.05), and more obvious attenuation was observed with a combination thereof (all p<0.01).
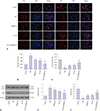
Propofol alleviates H/R-induced autophagy
To investigate whether propofol pretreatment attenuates the activation of H/R-induced autophagy, we also tested the levels of autophagy markers (LC3 and p62) via immunofluorescence and Western blot. As shown in Fig. 6A–C, the fluorescence intensity of anti-LC3II antibody was higher in the H/R group than that in the no-H/R control group (p<0.01), suggesting elevated levels of autophagy. This alteration was partially reversed by 50 µM propofol or 10 µM SP600125 (50 µM propofol versus H/R, p<0.01; 10 µM SP600125 vs. H/R, p<0.01). Like our findings with apoptosis, the combination of propofol and SP600125 provided greater alleviation against H/R-induced autophagy (p<0.01). In a similar vein, changes in the fluorescence intensity of anti-p62 antibody also suggested that propofol alleviated H/R induced autophagy (all p<0.05). Western blots further supported our hypothesis. Propofol administration significantly inhibited autophagy as demonstrated by decreased LC3II and increased P62 expression. Moreover, SP600125 strengthened the beneficial effect of propofol in HK2 cells exposed to hypoxia/reoxygenation injury (Fig. 6D–F; p<0.05).
DISCUSSION
To examine the effect of propofol on H/R injury and its underlying mechanism, an in vitro model was established by subjecting HK-2 cells to H/R insult in this study. Decades of experimental results have shown that PTCs play a vital role in the histopathology of renal injury, especially in I/R impairment.1112 After an acute ischemic insult, PTCs are the most sensitive cells to I/R injury, which is mainly due to physiological hypoxia in the medullary region and the high metabolic activity of PTCs.12 In recent years, numerous studies have used PTC models of H/R to investigate mechanisms of renal I/R injury or to search for drugs that lower the risk of I/R injury.1314 Several studies have shown that propofol is an excellent anti-inflammation and anti-oxidation agent, and may help to prevent and treat I/R injury,1516 although the exact mechanisms remain unclear. We investigated the protective effects and underlying mechanisms of propofol against H/R-induced injury in HK-2 cells in the absence or presence of the JNK inhibitor SP600125. H/R injury was evaluated by measuring cell viability, LDH leakage, and apoptotic index. Studies have indicated that the clinically relevant concentrations of propofol were approximately 11–56 µM.1718 Thus, the concentration range of 10–100 µM was used in this study, which made our results more clinically relevant. Our present study found a dose-dependent increase in cell viability, as well as dose-dependent decreases in LDH release and cell apoptosis, upon propofol treatment. All these indicate that propofol has a definite protective effect against H/R injury in HK-2 cells. Li, et al.16 reported that propofol prevented H/R injury in rat renal tubular epithelial cell line (NRK-52E cells), similar to our results. Compared with 50 µM of propofol, 100 µM offered no extra protection against H/R injury.
JNK is involved in cell apoptosis; however, it also acts synergistically with JAK/STAT, NF-κB, and other signaling molecules to promote cancer cell survival.19 Studies have reported that JNK activation after H/R is excessively increased in human brain vascular smooth muscle cells,20 H9c2 cardiomyocytes,21 and HK-2 cells.22 In the present study, we selected the specific JNK inhibitor SP600125 to ascertain the effect of propofol against H/R injury. Our results showed that propofol significantly attenuates H/R-induced cell apoptosis and JNK activation. Similar effects were shown by treatment with JNK inhibitor SP600125. These results indicated that the protective effects of propofol against H/R injury may be related to JNK inhibition. An interesting finding in our study was that the beneficial effects of the combined use of propofol and SP600125 were more obvious than use of only one them. These indicate their combination has better superimposed protective effects.
Autophagy is a critical mechanism for cellular homeostasis and survival in response to renal I/R injury. However, it is believed to be a double-edged sword in cell injury. On the one hand, physiologic autophagy is involved in degradation of harmful proteins and damaged organelles to prevent the accumulation of harmful substances, which limits the transmission of harmful signaling. On the other hand, excessive autophagy will cause irreversible injury and transform cells to autophagic cell death. To ascertain the role of autophagy in propofol-induced protection, HK-2 cells were pretreated with propofol in the absence or presence of SP600125 prior to H/R injury. We showed that autophagy was significantly increased after H/R as indicated by a significant increase in LC3II protein levels and a decrease in p62 protein levels, and propofol attenuated these changes, which were further alleviated by a combination of propofol and SP600125. Likewise, Xie, et al.23 reported that autophagosomes and autolysosomes were significant increased at 2 h after reoxygenation in HK-2 cells. According to previous reports, inhibition of the JNK pathway by SP600125 could block autophagy activation in cancer cells.24 We showed here that SP600125 attenuated H/R-triggered autophagy activation. Although the exact mechanism by which propofol-inhibited autophagy protects from H/R injury is unclear, our results showed that it is likely that propofol inhibits excessive autophagy during the injury partly through JNK pathway. It was confirmed that some situations, including overdigestion of cytoplasmic contents due to excessive autophagy, might result in autophagic cell death.25 Recent studies have reported that propofol antagonizes H/R-triggered neuronal injury via inhibition of autophagy.7 Other studies have stated that propofol postconditioning protects H9c2 cells from H/R injury by inducing autophagy via JNK pathway.26 This discrepancy could result from differences in the degree of autophagy activation or test subjects.
More should be done in future studies to verify the protective effect of propofol on H/R injury particularly with in vivo assay. Also, reoxygenation time may affect the expression levels of related proteins after H/R injury,14 and in this study, we harvested the cells only at 2 h after reoxygenation due to limited resources. Therefore, it would be informative to measure the effects of propofol against H/R injury at different reoxygenation times. Despite the limitations, our study has demonstrated unambiguously that propofol helps to protect HK-2 cells against H/R injury by alleviating apoptosis and autophagy, and suggests that the protective effects are derived from its inhibition of JNK activation.




 XML Download
XML Download