

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
According to the statistics, it has been reported that there were more than 14000 ovarian cancer-related deaths and 20000 new cases in 2018 in the United States.1 Although surgery and chemotherapy for ovarian cancer have greatly improved in recent years,2 overall survival remains less than satisfactory. After initial chemotherapy therapeutics, patients with advanced ovarian cancer often experience recurrence and die from chemoresistance disease.3 Tumor invasion, metastasis, and multidrug resistance have been found to be factors affecting tumor recurrence in ovarian cancer patients.4
The development of chemoresistance is becoming a major barrier to the treatment of cancer patients.5 Ovarian cancer patients can show poor responses to current treatment with cisplatin, a chemotherapeutic agent for long-term clinical application. It is imperative to investigate an effective therapeutic target than could enhance sensitivity to cisplatin and to elucidate related molecular pathways in order to develop promising therapy strategies for ovarian cancer with cisplatin resistance. Steg, et al.6 showed that ovarian cancer stem cells are involved in chemoresistance, metastasis, and tumor recurrence. Ricci, et al.7 reported that epithelial-mesenchymal transition (EMT) in ovarian cancer cells was associated with chemoresistance. Migration, attachment to mesothelial cells, and invasion into subperitoneal tissue were found to be involved in the formation of metastasis in peritoneal dissemination of gastric cancer.8 Chaffer and Weinberg9 also reported that EMT can modify the metastatic ability of cancer cells. Singh and Settleman10 agreed with the above conclusions that EMT of tumor cells increases metastasis and further indicated that it contributes to drug resistance. In addition, serial analyses of gene expression profiling have revealed that many EMT-related transcription factors are elevated in cisplatin-resistant cells.11
MiRNAs, consisting of 21–25 nucleotides, function as guide molecules in RNA silencing.12 Research has indicated that miRNA is associated with ovarian cancer drug resistance and can act as prognostic tools for assessing clinical outcomes.13 Furthermore, increasing evidence has shown that miR-338-3p acts as a tumor suppressor blocking the growth of thyroid cell by the targeting protein kinase Bγ.14 MiR-338-3p could inhibit proliferation, migration, invasion, and EMT in osteosarcoma cells by targeting activator of 90 kDa heat shock protein ATPase homolog 1.15 Meanwhile, Sun, et al.16 showed that miR-338-3p functions as a tumor inhibitor in gastric cancer by targeting protein-tyrosine phosphatase 1B. Consistent with the above findings, Liu, et al.17 demonstrated that miR-338-3p acts as a suppressor of epithelial ovarian cancer progression, although the mechanisms of drug resistance in cancer are poorly understood. In this study, we investigated the expression of miR-338-3p in cisplatin-resistant ovarian cancer tissues and cells. Functional and mechanism analyses were conducted to further clarify the relationships among miR-338-3p, WNT2B, and resistance to cisplatin in ovarian cancer.
MATERIALS AND METHODS
Clinical samples
In total, 54 frozen human primary ovarian tumor samples were divided into two groups according to their responses to the cisplatin: 24 treatment-responsive patients and 30 treatment-resistant patients, as well as normal non-tumors tissues, were collected from The First People's Hospital of Lianyungang. Biopsy samples were promptly frozen in liquid nitrogen and maintained at −80℃ until ready for further use. This study was approved by the Ethical Committee of The First People's Hospital of Lianyungang, and written informed consent was obtained from all participants before surgery. All biopsy samples used in this study were histopathologically examined by two independent pathologists.
Cells culture and transfection
SKOV3 cells (human ovarian carcinoma cell line) were purchased from the American Type Culture Collection (Manassas, VA, USA), and A2780 (human ovarian carcinoma cell line) and IOSE-80 (human ovarian epithelial cell line) cells were obtained from the Cancer Research Institute, Southern Medical University (Guangzhou, China). Cells were cultured in RPMI-1640 medium (GIBCO BRL, Grand Island, NY, USA) with 10% (v/v) fetal bovine serum (GIBCO BRL) in a 5% CO2 atmosphere at 37℃. Matched cisplatin-resistant ovarian cancer cells, named SKOV3/DDP and A2780/DDP cells, were produced from the parental cell lines and cultured with escalating concentrations of cisplatin as previously described.18 Overexpressed plasmids of miR-338-3p (miR-338-3p) and its negative control miR-NC, specific short hairpin RNA (shRNA) against WNT2B (sh-WNT2B) and its negative control sh-NC, and overexpressed plasmids of WNT2B (WNT2B) and its negative control pcDNA were purchased from RiboBio (Guangzhou, China). The above oligonucleotides or plasmids were transfected into ovarian cancer cell lines utilizing Lipofectamine 2000 reagent (Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturer's instructions.
RNA isolation and real-time quantitative polymerase chain reaction
Total RNA from cells and tissues was extracted with TRIzol reagent (Thermo Fisher Scientific). The quality of samples was regard as good when OD260 nm/OD280 nm values were greater than 1.8. The Prime Script RT Reagent kit (Takara, Dalian, China) and microRNA Reverse Transcription kit (Thermo Fisher Scientific) were employed to reverse transcribe RNA into complementary DNA. The expression levels of miR-338-3p and WNT2B were analyzed using SYBR Premix Ex Taq II (Takara) for real-time quantitative polymerase chain reaction (RT-qPCR) on an ABI 7500 HT system (Applied Biosystems, Foster City, CA, USA). Relatively quantitative analysis was performed for mean values using the 2−ΔΔCt method. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) or endogenous small nuclear RNA U6 served as an internal control.
The following primers were utilized: miR-338-3p (forward, 5′-ATCCAGTGCGTGTCGTGG-3′; reverse, 5′-TGCTTCCAGC ATCAGTGAT-3′); WNT2B (forward, 5′-ATTTCCCGCTCTGG AGATTT-3′; reverse, 5′-AAGCTGGTGCAAAGGAAAGA-3′); GAPDH (forward, 5′-TCCCATCACCATCTTCCAGG-3′; reverse, 5′-GATGACCCTTTTGGCTCCC-3′); and U6 (forward, 5′-CTCGCTTCGGCAGCACA-3′; reverse, 5′-AACGCTTCACG AATTTGCGT-3′).
MTT assay
Cell viability was detected with 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyl-2H-tetrazol-3-ium bromide (MTT) assay. Firstly, cells were seed onto 96-well plates at a density of 5.0×103 cells per well and treated with various doses of cisplatin for 48 h. After this, 20 µL of 5 mg/mL MTT (Sigma-Aldrich Co., St. Louis, MO, USA) was added to each well and incubated at 37℃ for another 4 h. Formazan crystals were then dissolved with dimethyl sulfoxide (DMSO). Absorbance at a wavelength of 490 nm was measured using a microplate reader (Applied Biosystems). Relative survival curves were drawn to determine IC50 values for the dose of cisplatin causing 50% inhibition of growth among ovarian cancer cells.
Flow cytometry analysis of apoptosis
SKOV3/DDP and A2780/DDP cells (3.5×105/well) after transfection were seeded into a 6-well plate treated with various doses of cisplatin for 48 h. 500 µL of binding buffer was then added to each well. Subsequently, 5 µL of propodeum iodide (Thermo Fisher Scientific) and 5 µL of Annexin V-Fluorescein isothiocyanate (Thermo Fisher Scientific) were added at room temperature for 15 min. The ratio of apoptotic cells was detected using a flow cytometer (Applied Biosystems).
Western blot assay
Total protein was subjected to sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene fluoride (PVDF) membranes (Millipore, Billerica, MA, USA). Subsequently, membranes were blocked with 5% nonfat milk before incubation with 1:2000 dilution primary antibody at 4℃ overnight, and then the membranes were incubated with secondary antibody with horseradish peroxidase for 2 h at a 1:5000 dilution. Finally, the signal intensities of antigen-antibody complexes were visualized using chemiluminescence.
Cell migration and invasion assay
For invasion assay, cells were serum starved for 24 h prior to use and were placed into 24-well plates with a polycarbonate membrane with 8-µm pores (Thermo Fisher Scientific) and Matrigel (Thermo Fisher Scientific) at 5×104 cells/well. Lower chambers containing fetal bovine serum were used to induce cells through membranes. Plates were incubated for 24 h at 37℃ in 5% CO2. After 24 h, the average number of invaded cells was calculated in five random fields of view under a microscope (Applied Biosystems) to produce a diagram of invasion cell numbers. Similarly, migration assays were conducted as described above, except the use of Matrigel was omitted.
Dual-luciferase reporter assay
The 3′UTR segments of WNT2B, which were predicted to interact with miR-338-3p using starBase2.0 (http://starbase.sysu.edu.cn/starbase2/index.php), were amplified by PCR and cloned downstream of the Renilla luciferase stop codon in pGL3 vector (Thermo Fisher Scientific), named WT or MUT reporters, respectively. Ovarian cancer cells were infected with reporter vectors using Lipofectamine 2000 (Thermo Fisher Scientific) in accordance with the provided instruction manual. Cells were harvested at 24-h post-transfection, and firefly or Renilla fluorescence signals were read using the Dual-Luciferase Reporter Assay System (Thermo Fisher Scientific). Renilla luciferase served as an internal control. Three independent experiments were performed.
In vivo experiment
In total, 21 male BALB/c nude mice were purchased from Shanghai Experimental Animal Center (Shanghai, China) and fed according to institutional guidelines. SKOV3/DDP cells transfected with miR-NC or miR-338-3p were subcutaneously injected into the right backs of the nude mice. The mice were treated with PBS or cisplatin (5.0 mg/kg) with intraperitoneally injected for one cycle of three days at 7-d post-transfection. Tumor growth was monitored every week, and size was calculated using V=1/2×ab2 [maximum (a) and minimum (b) length of the tumor]. At 35 d after injection, mice were sacrificed, and tumor samples were weighed and analyzed by Western blot and RT-qPCR assays. Animal experiments were approved by the Institutional Animal Care and Use Committee of The First People's Hospital of Lianyungang.
Statistical analysis
All statistical analyses were conducted with GraphPad Prism 7 (GraphPad Inc, La Jolla, CA, USA), and results are expressed as mean±standard deviation. p values less than 0.05 were regard as statistically significant. Differences between two groups were analyzed by Student's t-test. One-way analysis of variance was used to estimate differences among three or more groups.
RESULTS
MiR-338-3p expression is low in cisplatin-resistant ovarian cancer tissues and cells
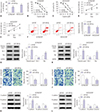
To begin, RT-qPCR was performed to detect the expression of miR-338-3p in normal tissues and ovarian cancer tissues (DDP-sensitive/DDP-resistance), as well as in IOSE-80, A2780, and SKOV3 cells and cisplatin-resistant cells (A2780/DDP and SKOV3/DDP cells). As presented in Fig. 1A and B, the data showed that miR-338-3p was significantly downregulated in ovarian cancer tissues and cells, compared with matched controls. Moreover, a marked decrease in miR-338-3p expression was observed in cisplatin-resistant tissues and cells, compared with their cisplatin-sensitive counterparts. Kaplan-Meier analysis with log-rank test further showed that low expression levels of miR-338-3p led to a lower survival rate of ovarian cancer patients, while high expression of miR-338-3p facilitated a higher survival rate (Fig. 1C). We concluded that expression of miR-338-3p is associated with cisplatin resistance in ovarian cancer patients.
Overexpression of miR-338-3p enhances sensitivity to cisplatin in ovarian cancer cells
Gain-of-function experiments confirmed that the expression levels of miR-338-3p were increased in cisplatin-resistant ovarian cancer cells transfected with miR-338-3p mimic (Fig. 2A). The IC50 values of cisplatin in A2780/DDP and SKOV3/DDP cells transfected with miR-338-3p were lower than those in miR-NC groups (Fig. 2B and C). To further explore the proliferative ability of A2780/DDP and SKOV3/DDP cells transfected with miR-NC or miR-338-3p, MTT assay indicated that high expression of miR-338-3p repressed the proliferation of ovarian cancer cells (Fig. 2D and E). In addition, overexpression of miR-338-3p elicited a higher apoptosis rate in miR-338-3p groups than that in miR-NC groups (Fig. 2F). Changes in apoptosis-related proteins were consistent with changes in apoptotic rate: upregulation of miR-338-3p promoted the expressions of cleaved-caspase-3 and Bax, but inhibited the expression of Bcl-2 (Fig. 2G and H). Transwell migration and invasion assays showed that upregulation of miR-338-3p inhibited migration and invasion of A2780/DDP and SKOV3/DDP cells (Fig. 2I and J). Investigation of the effects of miR-338-3p overexpression on the protein levels of E-cadherin, N-cadherin, and Vimentin by Western blot assay showed that protein levels of N-cadherin and Vimentin were remarkably reduced, while E-cadherin levels increased in response to transfection of miR-338-3p into A2780/DDP and SKOV3/DDP cells (Fig. 2K and L). Furthermore, levels of alpha smooth muscle actin (α-SMA) and fibronectin were mitigated in A2780/DDP and SKOV3/DDP cells transfected with miR-338-3p mimic (Supplementary Fig. 1A, only online). Collectively, miR-338-3p enhanced the sensitivity of ovarian cancer cells to cisplatin.
MiR-338-3p negatively targets WNT2B
WNT2B and its mutant sites are shown in Fig. 3A. Additionally, the results of dual-luciferase reporter assay revealed that elevated expression of miR-338-3p significantly repressed the luciferase activity of WNT2B 3′UTR-WT reporter, but not that of WNT2B 3′UTR-MUT reporter in A2780/DDP and SKOV3/DDP cells (Fig. 3B and C). To further clarify the expression levels of WNT2B in ovarian cancer cells and tissues, the protein and mRNA levels of WNT2B were assessed in treatment-responsive patient tissues or cells (A2780 and SKOV3) and treatment-resistant patient tissues and cells (A2780/DDP and SKOV3/DDP), as well as matched normal controls, utilizing RT-qPCR and Western blot assays. We discovered that the expression levels of WNT2B in cisplatin-resistant ovarian cancer cells (A2780/DDP and SKOV3/DDP) or tissues were markedly increased (Fig. 3D–F). Furthermore, a negative correlation between WNT2B and miR-338-3p was confirmed by RT-qPCR assay (Fig. 3G). Subsequently, we assessed the protein and mRNA levels of WNT2B in A2780/DDP and SKOV3/DDP cells introduced with miR-NC, miR-338-3p, miR-338-3p+pcDNA, or miR-338-3p+WNT2B. As shown in Fig. 3H–K, miR-338-3p overexpression markedly inhibited the expression levels of WNT2B at both the protein and mRNA level; meanwhile, reintroducing WNT2B could attenuate miR-338-3p-induced reductions in WNT2B. Taken together, these data demonstrated that miR-338-3p targets WNT2B expression.
WNT2B knockdown sensitizes A2780/DDP and SKOV3/DDP cells to cisplatin
RT-qPCR and Western blot assays indicated that WNT2B expression is reduced in A2780/DDP and SKOV3/DDP cells transfected with sh-WNT2B in comparison with matched control (Fig. 4A and B). Cells were treated with different doses of cisplatin for 48 h, and then IC50 values were detected by MTT assay. The results showed that WNT2B knockdown significantly decreased cisplatin resistance among A2780/DDP and SKOV3-8/DDP cells (Fig. 4C and D). Cell viability determination results showed that WNT2B knockdown significantly inhibited proliferation of A2780/DDP and SKOV3/DDP cells (Fig. 4E and F). Flow cytometry assay showed that apoptosis rates increased in SA2780/DDP and SKOV3/DDP cells after treatment with sh-WNT2B (Fig. 4G). Assessment of apoptosis-related proteins by Western blot assay revealed that decreased expression of WNT2B promotes the expression of cleaved-caspase-3 and Bax, but impedes the expression of Bcl-2 (Fig. 4H and I). Migration and invasion assays were carried out to evaluate motility ability of ovarian cancer cells. The results suggested that downregulation of WNT2B suppresses migration and invasion of cisplatin-resistance cells (Fig. 4J and K). Analogously, WNT2B silencing suppressed EMT processes in ovarian cancer cells by reducing N-cadherin and Vimentin expression, while promoting E-cadherin (Fig. 4L and M). Meanwhile, consistent with N-cadherin and Vimentin, α-SMA and fibronectin were inhibited by sh-WNT2B in A2780/DDP and SKOV3/DDP cells (Supplementary Fig. 1B, only online). Collectively, these data suggested that WNT2B knockdown weakens resistance to cisplatin among A2780/DDP and SKOV3/DDP cells.
MiR-338-3p impedes cisplatin resistance in ovarian cancer cells by downregulating WNT2B expression
A2780/DDP and SKOV3-8/DDP were transfected with miR-NC, miR-338-3p, miR-338-3p+pcDNA, or miR-338-3p+WNT2B. The transfected A2780/DDP and SKOV3/DDP cells were treated with various doses of cisplatin for 48 h, and IC50 values of cisplatin were measured by MTT assay. The results thereof showed that gain of WNT2B overturned upregulation of miR-142-5p-mediated resistance to cisplatin in ovarian cancer cells (Fig. 5A and B). Furthermore, we also found that greater expression of WNT2B reversed the reduced viability in A2780/DDP and SKOV3/DDP cells caused by miR-338-3p overexpression (Fig. 5C and D). As shown in Fig. 5E, upregulation of miR-338-3p resulted in a promotion of apoptosis in A2780/DDP and SKOV3/DDP cells, which was suppressed by overexpression of WNT2B, as demonstrated by flow cytometry analysis. Western blot assay was conducted to detect the protein expression levels of transfected cells and indicated that miR-338-3p overexpression markedly suppressed anti-apoptosis protein levels (Bcl-2) and enhanced pro-apoptosis protein levels (cleaved-caspase-3 and Bax). Reversal of WNT2B expression abolished the observed miR-338-3p-induced effects (Fig. 5F and G). Consistent with proliferation, overexpression of WNT2B abolished inhibition effects of miR-338-3p on migration and invasion in resistant ovarian cancer cells (Fig. 5H and I). Analogously, the protein levels of N-cadherin and Vimentin were apparently reduced, while E-cadherin was enhanced, in the miR-338-3p overexpression group, although upregulation of WNT2B overturned this effect (Fig. 5J and K). In addition, repression of α-SMA and fibronectin by treatment with miR-338-3p were abated in A2780/DDP and SKOV3/DDP cells by combining with WNT2B (Supplementary Fig. 1C, only online). Taken together, we determined that miR-338-3p directly targets WNT2B to reduce resistance in ovarian cancer cells.
Elevated miR-338-3p suppresses tumor growth in vivo
As presented in Fig. 6A and B, cisplatin treatment significantly decreased tumor volume and weight, while miR-338-3p+cisplatin treatment led to more obvious inhibition effects on the volumes and weights of tumors, compared to other groups. The expression levels of miR-338-3p were markedly higher in tumor tissues of the emiR-338-3p+cisplatin group than the other groups (cisplatin+miR-NC or miR-NC group); moreover, mRNA expression levels of WNT2B decreased in tumor tissues from the miR-338-3p+cisplatin group, compared with other groups (cisplatin+miR-NC or in miR-NC group) (Fig. 6C). In addition, overexpression of miR-338-3p induced a decrease in protein expression of WNT2B in tumor tissues (Fig. 6D).These results demonstrated that miR-338-3p represses tumor growth in vivo by affecting WNT2B expression.
DISCUSSION
In the present study, we found the expression levels of miR-338-3p to be associated with chemoresistance to cisplatin in ovarian cancer patients. Functional and mechanical experiments further revealed that miR-338-3p enhances sensitivity to cisplatin via downregulating WNT2B in ovarian cancer cells.
Previous study have shown that miRNA-630 inhibitor has a similar function as miR-338-3p and that suppression of miR-630 could sensitize SKOV3-TR cells to paclitaxel by increasing apoptosis.19 Analogously, miRNA-574-3p could target epidermal growth factor receptor to sensitize ovarian cancer cells to paclitaxel and cisplatin.20 Li, et al.21 indicated that miR-142-5p combined with cisplatin treatment could increase apoptosis in ovarian cancer cells by regualating apoptotic-associated genes expression. In addition, enforced expression of let-7 has bee found to enhance sensitivity to Taxol or vinblastine in ovarian cancer cells.22 Our data further showed that overexpression of miR-338-3p enhances sensitivity to cisplatin among ovarian cancer cells by regulating proliferation, apoptosis, and EMT process. Meanwhile, Zhang, et al.23 found that miR-338-3p regulates metabolism in ovarian cancer cell by inhibiting pyruvate kinase M2 expression. In the present study, bioinformatics analysis and dual-luciferase reporter assay were conducted and confirmed that miR-338-3p had putative binding sites on the 3′UTR of WNT2B: While WNT2B, WNT5A, WNT5B, and WNT9B all had putative binding sites with miR-338-3p, only co-transfection wild-type WNT2B and miR-338-3p showed reduced luciferase activity, as presented in Supplementary Fig. 2 (only online). Thus, the function of WNT2B was mainly explored in subsequence experiments. In view of the high expression of WNT2B in cisplatin resistance cells, the function of WNT2B knockdown in ovarian cancer cells was further explored. WNT2B plays a key role in Wnt/β-catenin signal pathway by regulating the proliferation, apoptosis, and EMT of cells. Jiang and Ma24 demonstrated that the expression of miR-324-3p in polycystic ovary syndrome rats is remarkably downregulated and confirmed that overexpression of miR-324-3p represses proliferation and induces the apoptosis of granulosa cells by targeting of WNT2B. Moreover, other have found that miR-185-3p regulates radio-resistance of nasopharyngeal carcinoma via targeting WNT2B in vitro.25 Furthermore, HOXD cluster antisense RNA 1 promoted proliferation, invasion, and EMT of epithelial ovarian cancer cells has been found to be affected by Wnt/β-catenin signaling.26 Altogether, the Wnt/β-catenin pathway appears to be involved in EMT as a major contributor to the tumorigenesis of ovarian cancer.27 In our research, the overexpression of miR-338-3p suppressed EMT by downregulating N-cadherin and Vimentin expression and by increasing E-cadherin. Importantly, upregulation of WNT2B overturned the effects of miR-338-3p overexpression on EMT. In agreement with our data, a review conducted by Du and Shim28 confirmed that EMT plays an important role in metastasis and drug resistance among cancer cells. E-cadherin, N-cadherin, and Vimentin are considered EMT-associated marker proteins. Research indicated that E-cadherin is an important molecule for cell-cell adhesion and functions as a biomarker of EMT. Indeed, low expression of E-cadherin has been shown to facilitate separation of cancer cells from the serosal surface.29
Here, we explored the influence of miR-338-3p on cisplatin resistance in ovarian cancer and confirmed that the expression of miR-338-3p is positively correlated with 5-year survival rate in ovarian cancer patients. We also showed that expression of miR-338-3p enhances the sensitivity of ovarian cancer cells to cisplatin in vivo and in vitro.
Regrettably, our experiment did not conduct a comprehensive study on drug metabolism-related proteins. Our results demonstrated that abundant expression of miR-338-3p suppresses resistance to cisplatin in ovarian cancer cells by affecting WNT2B. In the future research, we will focus on whether the impact effect of miR-338-3p/WNT2B on cisplatin resistance is associated with drug metabolism signaling pathways. Knowing the molecular factors involved in drug resistance may help us to develop new strategies for chemotherapy and reduce the relapse rate of ovarian cancer.
In conclusion, our data suggest that miR-338-3p is involved in cisplatin resistance in ovarian cancer. MiR-338-3p appears to act as a therapeutic target through which to enhance sensitivity of ovarian cancer cells to cisplatin by targeting WNT2B.





 XML Download
XML Download