

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Liver fibrosis ultimately leads to adverse consequences, such as liver cirrhosis, portal hypertension, hepatocellular carcinoma, and hepatic failure, making it a major cause of mortality and morbidity worldwide.1 Advanced liver fibrosis exhibits the accumulated extracellular matrix (ECM) constituents, such as collagen, proteoglycan, and laminin, in the liver parenchyma.23 In response to chronic liver injuries, macrophages and platelets release transforming growth factor-β (TGF-β) that is a potent activator of hepatic stellate cells (HSCs).3 Subsequently, activated HSCs transdifferentiate into myofibroblastic cells that then participate in synthesis and reorganization of connective tissue, becoming a principal producer of ECM components.456 However, the detailed molecular mechanisms underlying liver fibrosis are very intricate and still largely unknown.
MicroRNA (miRNA) refers to a small non-coding RNA molecule containing about 20 to 24 nucleotides that function in RNA silencing and post-transcriptional regulation of gene expression. MicroRNAs are incorporated into an RNA-induced silencing complex via complementary base-pairing within mRNA molecules. Consequently, these mRNA molecules are silenced, thereafter moving to the targeted site wherein they play a role in translational inhibition or destabilization of the target mRNA molecule. MicroRNAs act as essential players in a number of diseases, including liver fibrosis789101112; they participate in the process of liver fibrosis, by either promoting or inhibiting it.131415
We have performed several in vitro and in vivo experiments to find a way to ameliorate liver diseases using secretome.161718192021 Secretome herein refers to the total set of molecules secreted or surface-shed by stem cells. The reasons why we used secretome instead of stem cells include 1) the instability of stem cells, such as the potential of malignant transformation, 2) proven mechanism of action of stem cells that is secretome-mediated, and 3) the easy accessibility of secretome. As a means of enhancing the therapeutic potential of secretome, we had attained the secretome from adipose-derived stem cells (ASCs) that had been transfected with miR-214.2223242526 In general, miR-214 is known to exhibit an antifibrotic effect by inhibiting connective tissue growth factor (CCN2) that promotes the activation of HSCs.22 However, several studies also reported that miR-214 promotes HSCs, thereby prompting liver cirrhosis.2224 The pathophysiological roles of miR-214 in liver fibrosis remain largely unknown. In this study, we hypothesized that the secretome released from miR-214-transfected ASCs (miR-214-secretome) exhibits higher antifibrotic potential than that of naïve secretome in the mouse model of liver fibrosis.
METHODS
Preparation of cells
The lipoaspirated fat was digested by 0.1% collagenase (Sigma-Aldrich, St. Louis, MO, USA) in saline and collected after centrifugation. Cells were plated into culture flask in low-glucose Dulbecco's Modified Eagle's Medium (DMEM; Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% FBS (Thermo), 100 U/mL of penicillin (Thermo), and 0.1 mg/mL of streptomycin (Thermo). ASCs were incubated at 37°C in humidified chamber containing 5% carbon dioxide and medium was changed every 3 days.
Transfection of miR-214 into ASCs
ASCs were transfected with miRNA-214 (miR-214; Exiqon, Germatown, MD, USA) using Lipofectamine RNAiMAX Reagent (Thermo) in medium per well. After 72 hours of transfection, the cells were morphologically observed by the inverted microscope. The cell numbers were counted with an automatic cell counter (Countess; Invitrogen, San Diego, CA, USA) using trypan blue solution. Transfected cells were processed for cell phenotyping or differentiated into adipogenic and osteogenic progenitor cells. For the characterization of cells, the immunophenotypes of naïve or transfected ASCs were determined by flow cytometry analysis (Cytomics FC500 flow cytometer; Beckman Coulter, Fullerton, CA, USA) using FITC-conjugated CD31, CD45, and CD73 antibodies and PE-CD105 antibodies (BD Pharmingen, San Jose, CA, USA). Isotype controls were performed with antibodies against IgG for samples. For the identification of multilineage differentiation potential, ASCs were cultured in STEMPRO® differentiation media (adipogenesis and osteogenesis differentiation kits; Gibco, Carlsbad, CA, USA) for 3–4 weeks. The cells were stained with Oil Red O or Alizarin Red for the identification of lipid vesicles (in adipocytes) and calcium deposits (in osteocytes), respectively. Nucleus visualization was accomplished with DAPI counterstaining. The cells were analyzed with a FASC Canto II (BD Biosciences, San Jose, CA, USA).
Design of animal study
We used five-week male BALB/c mice (Orient Bio, Seongnam, Korea) in this study. We then compared the effects of the MCM in an in vivo model of thioacetamide (TAA)-induced hepatic fibrosis model. The in vivo model was generated by subcutaneous injection of TAA (200 mg/kg, three times a week for 5 weeks) into the mice. The mice were intravenously infused with normal saline (0.1 mL of saline; n = 14), control secretome (0.1 mL equivalent of the 25-fold concentrated secretome obtained from 105ASCs after 24 hours culture in serum-free medium, n = 14), or miR-214-secretome (0.1 mL equivalent of the 25-fold concentrated serum-free medium obtained from 105ASCs transfected with miR-214, n = 14) once a week for 2 weeks, respectively.
Real-time quantitative PCR
Total cell RNA was extracted using TRIzol reagent (Invitrogen) according to the manufacturer's instructions. Reverse transcription was performed with 1 μg RNA using RT-Premix kit (TOYOBO, Osaka, Japan) according to the manufacturer's instructions. The primers used for SYBR Green real-time quantitative (q) PCR were as follows: TGF-β1, forward 5′-CCCCACTGATACGCCTGAGT-3′ and reverse 5′-AGCCCTGTATTCCGTCTCCTT-3′; MMP-2 forward 5′-CAGGGAATGAGTACTGGGTC-3′ and reverse 5′-ACTCCAGTTAAAGGCAGCAT-3′; α-SMA forward 5′-CCAGAGCAAGAGAGGGATCC-3′ and reverse 5′-TGTCGTCCCAGTTGGTGATG-3′; GAPDH, forward 5′-CGACTTCAACAGCAACTCCCACTCTTCC-3′ and reverse 5′-TGGGTGGTCCAGGGTTTCTTACTCCTT-3′. The reaction was performed using an Applied Biosystems Step one plus Real-Time PCR system (Thermo). After normalization to the GAPDH gene, the expression levels for each target gene were calculated using the comparative threshold cycle method. The data are presented as the mean ± standard deviation (SD) from three independent experiments.
Western blot analysis
Liver specimens obtained from mice were lysed using the EzRIPA Lysis kit (ATTO Corporation; Tokyo, Japan), and quantified by Bradford reagent (Bio-RadHercules, CA, USA). Proteins were visualized by western analysis using the following primary antibodies (1:1,000 dilution) at 4°C overnight and then with HRP-conjugated secondary antibodies (1:2,000 dilution) for 1 hour at 25°C. From Cell Signaling Technology (Beverly, MA, USA), we obtained primary antibodies against PCNA (proliferating cell nuclear antigen), TGF-β1 (transforming growth factor-β1), α-SMA (alpha-smooth muscle actin), TIMP-1 (Metallopeptidase inhibitor 1), MMP-2 (Matrix Metallopeptidase-2), β-actin, and horseradish peroxidase (HRP)-conjugated secondary antibody. Specific immune complexes were detected using the Western Blotting Plus Chemiluminescence Reagent (Millipore, Bedford, MA, USA).
Enzyme-linked immunosorbent assay (ELISA)
Blood samples were collected from each mouse, centrifuged for 10 minutes at 9,500 g, and serum was collected. The concentrations of mouse interleukin (IL)-6 and tumor necrosis factor (TNF)-α were measured by sandwich ELISA (Biolegend, San Diego, CA, USA) according to the manufacturer's instructions.
Special staining including immunohistochemistry
Masson's trichrome staining and Sirius red staining were performed using the Sirius red staining kit and Masson's trichrome staining kit (Polysciences, Warrington, PA, USA), respectively, according to the manufacturer's protocol. For immunohistochemistry (IHC) analysis, formalin-fixed, paraffin-embedded tissue sections were deparaffinized, rehydrated in an ethanol series and subjected to epitope retrieval using standard procedures. Antibodies against of α-SMA, superoxide dismutase (SOD), and albumin (all from Cell Signaling Technology, MA, USA) were used for IHC staining. The samples were then examined under a laser-scanning microscope (Eclipse TE300; Nikon, Tokyo, Japan) to analyze the expression of these antibodies.
Statistical analysis
All data were reported using means ± SD. Statistical analysis was performed using SPSS (Ver. 11.0; SPSS Inc., Chicago, IL, USA). Statistical comparison among groups was determined using Kruskal-Wallis test. Probability values of P < 0.05 were regarded as statistically significant.
Ethics statement
Human ASCs were obtained from lipoaspirated fat with informed consent of the volunteers. This research was approved by Institutional Review Board (IRB No. 700069-201407-BR-002-01) of Hurim BioCell Co. Ltd. (Seoul, Korea). Animal studies were carried out in compliance with the guidelines of the Institute for Laboratory Animal Research, Korea (IRB No: CUMC-2018-0175-01).
RESULTS
Determination of stability of miR-214-transfected ASCs
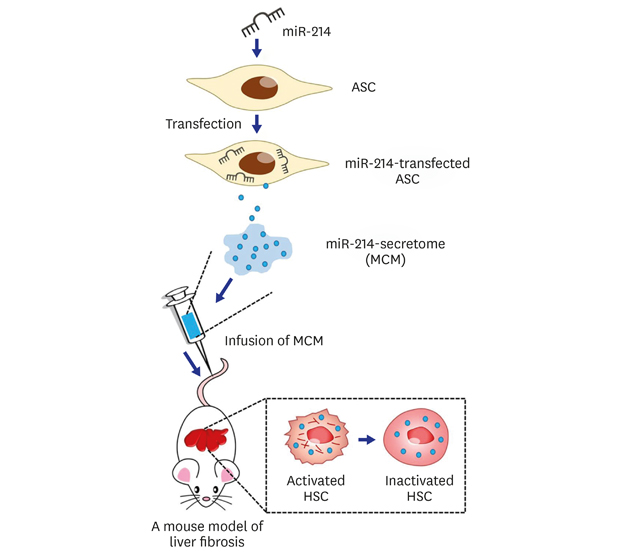
Fig. 1A shows the schematic illustration of this study. We obtained miR-214-secretome from conditioned media in which miR-214-transfected ASCs had been cultured for 48 hours. Subsequently, we planned to intravenously infuse the miR-214-secretome into the mice with liver fibrosis, and to determine the effects of miR-214-secretome on liver fibrosis. We first intended to determine the stability of miR-214-transfected ASCs. To achieve this goal, we determined surface marker expression and multilineage differentiation ability of the miR-214 transfection ASCs. Transfecting miR-214 into ASCs did not alter the gross morphology of cultured ASCs (Fig. 1B). Similar to non-transfected ASCs, the miR-214 transfection ASCs expressed mesenchymal stem cell markers (CD73 and CD105) and did not express hematopoietic stem cell markers (CD31 and CD45) (Fig. 1C). Finally, we successfully differentiated miR-214-transfected ASCs into adipocytes and osteocytes, demonstrating the preserved multilineage differentiation potential of ASCs following miR-214 transfection (Fig. 1D).
 | Fig. 1Determination of stability of miR-214-transfected ASCs. (A) A schematic illustration of study concept. microRNA-214 secretome is obtained from conditioned media in which miR214-transfected ASCs were cultured for 48 hours. Subsequently, we intravenously infused miR-214-secretome into mice with liver fibrosis, and determined the effects of miR-214-secretome on liver fibrosis. (B) Comparison of gross morphology between ASCs either or not transfected with miR-214. Transfecting miR-214 into ASCs did not alter the gross morphology of cultured ASCs. (C) Flow cytometry analysis of expressions of surface markers on ASCs transfected with miR-214. The miR-214-transfected ASCs were negative for CD31 and CD45 (hematopoietic stem cell markers) and positive for CD73 and CD105 (mesenchymal stem cell markers), similar to non-transfected ASCs. (D) Validation of preserved differentiation potential after transfecting ASCs with miR-214. Adipogenic (Left) and osteogenic (Right) differentiation of miR-214-transected ASCs was identified using Oil Red O and Alizarin red stains, respectively (Scale bars = 200 µm). Values are presented as mean ± standard deviation of three independent experiments.ASCs = adipose-derived stem cells, MCM = the secretome released from miR-214-transfected ASCs, HSC = hepatic stellate cell.
*P < 0.05.
|
Antifibrotic effects of the miR-214-secretome in mice with liver fibrosis
Here, we intended to determine antifibrotic effects of miR-214-secretome in the mice with liver cirrhosis (n = 21) as well as those with preserved liver function (n = 21). The mouse model of liver fibrosis was established by subcutaneous injection of TAA (200 mg/kg) three times a week for 5 weeks. Subsequently, the mice in each group were intravenously infused with normal saline (n = 14), control secretome (200 mg/kg; n = 14), or miR-214-secretome (200 mg/kg; n =14) once a week for 2 weeks. The mice were euthanized for obtaining the specimens on the seventh day post-infusion.
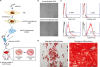
We first performed RT-PCR using the liver specimens for the determination of the RNA expression of fibrosis-related markers, such as α-SMA, TGF-β, and MMP-2 in each group (Fig. 2A). Although level of these markers increased significantly in the fibrotic liver specimens, the infusions of either naïve secretome or miR-214-secretome into the mice with liver fibrosis significantly reduced the RNA expression levels of these markers (P < 0.05). There was no significant difference in RNA expression levels of α-SMA and TGF-β between naïve secretome and miR-214-secretome groups. However, miR-214-secretome groups showed significantly reduced expression of MMP-2 compared with naïve secretome group (P < 0.05).
 | Fig. 2Antifibrotic effects of the miR-214-secretome in mice with liver fibrosis. The mice with or without liver fibrosis were intravenously infused with normal saline, control secretome, or miR-214-secretome two times a week for 1 week. (A) RT-PCR results demonstrating mRNA expression levels of α-SMA, TGF-β1, and MMP-2 in liver specimens of each group. The infusions of either naïve secretome (CM) or miR-214-secretome (MCM) into mice with liver fibrosis significantly reduced the RNA expression of these markers. miR-214-secretome groups showed significantly reduced expression of MMP-2 compared with naïve secretome group. (B) Results of western blot analysis demonstrating the effects of miR-214-secretome on the expression of various markers in mice with liver fibrosis. The infusion of the miR-214-secretome induced higher expression of PCNA, which reflects hepatocyte proliferation, as well as lower expression of fibrosis-related markers (α-SMA, TIMP-1, TGF-β1, and MMP-2) in the liver specimens than naïve secretome. Values are presented as mean ± standard deviation of three independent experiments.α-SMA = alpha-smooth muscle actin, CM = the secretome obtained from ASCs after 48-hour incubation, MCM = the secretome released from miR-214-transfected ASCs, TGF-β = transforming growth factor-β, MMP-2 = metalloproteinases-2, PCNA = proliferating cell nuclear antigen, TIMP-1 = tissue inhibitor of metalloproteinases-1.
*P < 0.05.
|
Next, we performed western blot analysis for comparing the levels of various markers in each group, including a proliferation marker (PCNA) and fibrosis-related markers (α-SMA, TIMP-1, TGF-β, and MMP-2) (Fig. 2B). Although the expression of PCNA was decreased in the fibrotic liver specimens, it was significantly increased in both secretome groups (P < 0.05). In addition, we found that the expression of fibrosis-related markers was significantly decreased after the infusions of either naïve secretome or miR-214-secretome into mice with liver fibrosis (P < 0.05). While comparing the two secretome groups, miR-214-secretome group significantly decreased the expression of all the fibrosis-related markers compared with that in naïve secretome group (P < 0.05).
Effects of miR-214-secretome on the liver enzymes and systemic inflammation
We performed serological analyses using blood samples on the seventh day post-infusion to compare the serum levels of AST and ALT in each group (Fig. 3A). Serum levels of AST and ALT were significantly elevated in mice with liver fibrosis; however, infusion of either kinds of secretome significantly reduced the elevated levels of AST and ALT (P < 0.05). Especially, miR-214-secretome significantly decreased serum levels of AST and ALT compared to that by naïve secretome (P < 0.05).
 | Fig. 3Effects of miR-214-secretome on liver enzymes and systemic inflammation. (A) Serological analysis using blood samples on seventh day post-infusion for the comparison of the serum levels of AST and ALT in each group. Infusion of either kinds of secretome significantly reduced the elevated levels of AST and ALT in the mice with liver fibrosis. Particularly, miR-214-secretome (MCM) significantly decreased serum levels of AST and ALT than naïve secretome (CM). (B) Results of ELISA using blood samples on seventh day post-infusion for the comparison of the serum levels of pro-inflammatory cytokines, such as IL-6 and TNF-α, in each group. In mice with liver fibrosis, secretome infusions significantly reduced serum levels of IL-6 and TNF-α than saline infusion. Of them, miR-214-secretome infusion induced the most significant reduction of these cytokines. Values are presented as mean ± standard deviation of three independent experiments.CM = the secretome obtained from ASCs after 48-hour incubation, MCM = the secretome released from miR-214-transfected ASCs, AST = aspartate transaminase, ALT = alanine transaminase, IL-6 = interleukin 6, TNF-α = tumor necrosis factor-α.
*P < 0.05.
|
We next performed ELISA using blood samples on the seventh day post-infusion to compare the serum levels of pro-inflammatory cytokines, such as IL-6 and TNF-α, in each group (Fig. 3B). In mice with liver fibrosis, secretome infusions significantly reduced serum levels of IL-6 and TNF-α compared with saline infusion (P < 0.05). Especially, miR-214-secretome infusion induced the most significant reduction of these cytokines (P < 0.05).
Effects of miR-214-secretome on the histology of the liver
To compare histological changes in each group, we performed various staining procedures reflecting the degree of inflammation and fibrosis, such as hematoxylin and eosin (HE), Masson's trichrome, and Sirius red staining. HE stains demonstrated that, although inflammation driven by TAA deranged the histological architecture of the liver, infusion of either kinds of secretome significantly recovered it (Fig. 4A). When comparing both secretome groups, miR-214 group exhibited higher degree of recovery from inflammation. Masson's trichrome and Sirius red stains demonstrated the superior antifibrotic activities of secretome infusions (P < 0.05). Between two secretome groups, miR-214-secretome group showed significantly higher amelioration of fibrosis compared to that by the control secretome group (P < 0.05) (Fig. 4B and C).
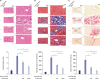 | Fig. 4Effects of miR-214-secretome on the histological characteristics of the liver specimens. (A) Top: H&E stains of the liver specimens in each group. Although inflammation driven by TAA deranged the histologic architectures of the liver, infusion of either kinds of secretomes significantly recovered it. Of the two secretome groups, histological improvement appeared to be more prominent in miR-214-secretome (MCM) group than in naïve secretome (CM) group. Bottom: The severity of liver injury was assessed using Suzuki classification. (B) Top: Masson's trichrome demonstrating the superior antifibrotic activities of secretome infusions. Of the two secretome groups, miR-214-secretome group showed significantly higher amelioration of fibrosis than naïve secretome group. Bottom: Relative densities of these markers quantified using ImageJ software. (C) Top: Sirius red stains demonstrating the superior antifibrotic activities of secretome infusions. Of the two secretome groups, miR-214-secretome group showed significantly higher amelioration of fibrosis than control secretome group. Bottom: Relative densities of these markers quantified using ImageJ software. Values are presented as mean ± standard deviation of three independent experiments.MCM = the secretome released from miR-214-transfected ASCs, CM = the secretome obtained from ASCs after 48-hour incubation, H&E = hematoxylin and eosin, TAA = thioacetamide.
*P < 0.05.
|
Effects of miR-214-secretome on immunohistochemical staining of the liver
For further elucidation of the role of miR-214-secretome, we performed the IHC staining of the liver specimens using antibodies reflecting fibrosis (α-SMA), antioxidant activity (SOD), and liver synthetic function (albumin). With respect to α-SMA IHC staining, infusion of miR-214-secretome significantly reduced the expression of α-SMA compared to saline infusion (P < 0.05) (Fig. 5A). With respect to SOD IHC, infusion of miR-214-secretome increased the expression of SOD compared to saline infusion (P < 0.05) (Fig. 5B). With respect to albumin IHC staining, infusion of miR-214-secretome significantly increased the expression of albumin compared to that by saline infusion (P < 0.05) (Fig. 5C).
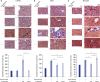 | Fig. 5Effects of miR-214-secretome on immunohistochemical staining of the liver. (A) Top: α-SMA IHC staining showing that infusion of miR-214-secretome (MCM) significantly reduced the expression of α-SMA compared to saline infusion. Bottom: Relative densities of these markers quantified using ImageJ software. (B) Top: SOD IHC staining showing that infusion of miR-214-secretome significantly increased the expression of SOD compared to that by saline infusion. Bottom: Relative densities of these markers quantified using ImageJ software. (C) Top: Albumin IHC staining showing that infusion of miR-214-secretome significantly increased the expression of albumin compared to saline infusion. Bottom: Relative densities of these markers quantified using ImageJ software. Values are presented as mean ± standard deviation of three independent experiments.α-SMA = alpha-smooth muscle actin, MCM = the secretome released from miR-214-transfected ASCs, CM = the secretome obtained from ASCs after 48-hour incubation, SOD = superoxide dismutase, IHC = immunohistochemistry.
*P < 0.05.
|
DISCUSSION
In this study, we intended to validate the superior antifibrotic potential of miR-214-secretome over naïve secretome in mice with liver fibrosis. We first generated the ASCs that had been transfected with antifibrotic miR-214, and extracted the miR214-secretome from conditioned media of the transfected ASCs through a series of ultrafiltrations. Liver fibrosis was ameliorated by intravenous infusion of miR-214-secretome into mice with liver fibrosis, which was demonstrated by significantly lower expression of fibrosis-related markers as well as lower fibrotic scores of special liver stains in the livers infused with miR-214-secretome than the livers infused with naïve secretome. The infusion of miR-214-secretome also led to the lesser local and systemic inflammation, higher expression of an antioxidant enzyme (SOD), and higher proliferative and synthetic functions of the liver. Taken together, we speculated that miR-214 transfection into ASCs led to reconditioning ASCs to release the secretome with higher antifibrotic and anti-inflammatory properties than those of naïve secretome.
Numerous investigators have turned their attention from stem cells to secretome after recognizing the limitations of stem cell research. Although still attractive due to its intrinsic advantages, stem cell research presents several challenges, including immune-mediated rejection, senescence-induced genetic instability or loss of function, and limited cell survival.26 Moreover, the major obstacle in the clinical application of stem cells is the possibility of malignant transformation.2728293031 Meanwhile, accumulating evidence has indicated that secretome released from stem cells exhibits similar potential as stem cells because the principal action of stem cells is mediated by the secretome.3233343536373839 Our previous experiments using secretome also demonstrated that secretome has similar, at least not inferior, therapeutic potential as stem cells in ameliorating hepatic failure.171819202140
Utilizing secretome from ASCs is one of the highly attractive ways of conducting stem cell research because adipose tissue itself is also considered as an endocrine organ. Mature adipocytes secrete a variety of proteins, collectively called adipokines, including adiponectin, leptin, plasminogen activated inhibitor 1, resistin, visceral adipose tissue-derived serine protease inhibitor (vaspin), and several proinflammatory cytokines.41424344454647 Moreover, ASCs are expected to be superior to mature adipocytes in terms of the number and concentration of secretory proteins. Lee et al.48 analyzed the components of human ASC secretome that had been induced by TNF-α using liquid chromatography–mass spectrometry. They identified 187 secretory proteins, including IL-6, IL-8, CXCL6 and MCP-1, cathepsin L, matrix metalloproteases, protease inhibitors, and pentraxin 3, all of which are known to be important mediators of inflammatory and immunomodulatory responses. They also revealed that the components and levels of these mediators vary according to the stimulatory duration and concentration of TNF-α.
There are two ways of applying secretome for a variety of disease models: without processing it and with processing it. It can lead to favorable and productive consequences to use secretome without processing. Moreover, the use of secretome without processing is worth considering because the components of secretome considerably vary according to its external environment that can be easily modified. We have attempted several ways of harnessing stem cells to release secretome with desired components, including hypoxic conditioning, chemical conditioning using lipopolysaccharides, and stimulating stem cells with pathogenic materials.171820 Here, we validated a novel way of stimulating ASCs to release secretome with desired components via transfecting ASCs with miRNAs that had been implicated in the pathogenesis of a targeted disease.
MicroRNA-214 is one of the representative antifibrotic miRNAs known to be a direct negative regulator of CCN2.22 CCN2 is a complex matricellular molecule that is produced downstream of TGF-β signaling and directly regulates the functions of activated HSCs, including mitogenesis, chemotaxis, adhesion, matrigenesis, and fibrogenesis.49 Therefore, CCN2 is considered as an essential regulator of TGF-β signaling during the fibrotic process. Quiescent HSCs exhibit depressed level of CCN2 because miR-214 inhibits CCN2 expression by its direct binding to the CCN2 3′ untranslated region (UTR).49 However, activated HSCs exhibit higher level of CCN2, which is related to lower level of miR-214, demonstrating reciprocal expressions of miR-214 and CCN2 during the progression of liver fibrosis.22
However, the role of miR-214 needs to be more clearly determined by further studies. In our experiments, for example, MCM showed significant antifibrotic effects on fibrotic liver, but not on normal liver, but increased fibrosis-related markers. For example, in our experiment, whereas MCM showed significant antifibrotic effects on fibrotic livers, it also increased the expression of fibrosis-related markers in the nonfibrotic livers, although statistically insignificant. This suggests that miR-214 may have a profibrotic effect in certain circumstances. Denby et al.50 reported that genetic deletion of miR-214 in mice significantly attenuated renal fibrosis induced by unilateral ureteral obstruction, suggestive of profibrotic effects of miR-214. In the heart, genetic deletion of miR-214 led to increased fibrosis through repression of sodium/calcium exchanger 1 (Ncx1), a key regulator of calcium influx.51 These findings suggest that the pathological role of miR-214 in fibrosis could be different according to organs and individual environment.
Until now, miR-214 had been investigated exclusively with respect to HSCs.2223242526 In this study, we first transfected miR-214 into ASCs, and investigated whether miR-214-transfected ASCs release the secretome with higher antifibrotic properties. Intravenous infusion of miR-214-secretome significantly reduced the expression of fibrosis-related markers, and reduced the serum levels of pro-inflammatory mediators in the in vivo model of liver fibrosis, demonstrating its superior antifibrotic and anti-inflammatory properties over naïve secretome. We thus speculated that miR-214 transfection into ASCs leads to reconditioning of ASCs to release the secretome with higher antifibrotic and anti-inflammatory properties than those of the naïve secretome.
We speculated that there are two ways of utilizing miR-214-secretome for clinical application. The first way is to deliberately refine the process of attaining miR-214-secretome for maximizing the antifibrotic efficiency of miR-214-secretome. The refinements include the determination of optimal concentration of miR-214, timing and duration of transfection and culturing, and whether or not to add supplementary physicochemical conditioning. The second way is to identify the causable factors among secretome components. Recently, a variety of studies have utilized proteomic approach to identify proteins the abundance levels of which are altered by a disease.525354 Mass spectrometry-based proteomic analysis now allows for measurement of the absolute or relative abundance of thousands of proteins simultaneously, representing a powerful analytic tool used in numerous clinical laboratories worldwide. Therefore, proteomics and mass spectrometry technologies could provide a promising approach for direct identification of specific therapeutic proteins in secretome. However, it is still questionable whether it is possible to identify a few therapeutic proteins because therapeutic effects of secretome could be attributed to the cumulative effect of rising and falling concentrations of various proteins constituting the secretome.
In conclusion, our study demonstrated that miR-214 transfection into ASCs reconditioned ASCs to release secretome with higher antifibrotic and anti-inflammatory properties. Specifically, liver fibrosis was ameliorated by intravenous infusion of miR-214-secretome into mice with liver fibrosis, which was demonstrated by significantly lower expression of fibrosis-related markers in the livers as well as lower fibrotic scores in the special stained livers compared with naïve secretome. The infusion of miR-214-secretome also led to the lower degree of local and systemic inflammation, higher expression of an antioxidant enzyme (SOD), and higher liver proliferative and synthetic functions. MicroRNA-214-secretome is thus expected to be a potential agent that can be used for overcoming liver fibrosis, if future studies can validate its safety and efficiency in preclinical experiments and clinical trials.
 XML Download
XML Download