

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
The prevalence of allergic diseases has increased steadily during the past 50 years,123 and multiple environmental, food, and innate immunological factors are implicated as the main causes of these disorders.4567 The recent marked and rapid increase in allergic diseases suggests that changes in the modern environment may be the underlying cause. Some of the suspected triggers are delivery mode, perinatal treatment with antimicrobials, and changes in infant feeding, all of which can influence gut microbiota composition.4689 The hygiene hypothesis has also been advanced and stresses the role of gut microbiota in the onset of allergic disorders.1011
A considerable number of studies have recently investigated the relationship between gut microbiota and allergic diseases in humans. Several of them reported that the development of an allergic disease correlates with insufficiency in gut microbiota compared to the healthy gut.812 A lower exposure to microbiota early in life can contribute to increased immune dysregulation and lead to a variety of diseases, including allergic, autoimmune, and metabolic disorders, as well as neoplastic diseases.913 A decreased level of short-chain fatty acids (SCFAs) and suppression of the innate immune response have also been reported as mechanisms that can affect the gut microbiota.81415 There is a constant life-long interaction between external stimuli and gut microbiota, and this interaction is important for maintaining homeostasis in the intestinal mucosal immune system.161718
The increasing use of antibiotics in modern-day medicine also has ramifications for gut intestinal microbiota. The administration of broad-spectrum antibiotics in early life is reported to increase susceptibility to Th17-mediated psoriasis and Th2-mediated atopic dermatitis (AD).192021 Moreover, an imbalance in gut bacterial homeostasis has been recognized as a crucial factor in the development of atopic disorders.122223 The postulated mechanism supposes that gut epithelial inflammation results from dysbiosis of the gut microbiota, which underlies chronic AD progression.12 To date, however, there has been no clear demonstration of an interaction between a gut microbiota imbalance and the progression of AD.
Most studies investigating the interaction between gut microbiota and human AD have employed clinical data from fecal analyses, which cannot fully serve to evaluate the effects on immunological mechanisms. Moreover, not enough is known of the mechanism involved in AD to confirm the influence of gut dysbiosis.
We have previously reported that oral probiotic administration can attenuate allergic inflammation in AD and allergic march in mouse models.1011 Our previous findings suggested that beneficial intestinal microbes can suppress allergic inflammation in AD. In our present study, we investigated whether gut dysbiosis caused by administration of an oral antibiotic cocktail aggravates allergic inflammation in a mouse model of AD and identified whether the underlying mechanism involves intestinal innate immune cells, metabolites, and the systemic immune response. We further evaluated the role of gut microbiota homeostasis in the development of AD as well as the associated mechanisms.
MATERIALS AND METHODS
AD mouse model
The AD mouse model was generated via ovalbumin (OVA) sensitization as previously described, with some modifications.24 Briefly, 6-week-old female mice were anesthetized and their backs were shaved. Skin injury was then induced by repeated tape stripping, after which 100 µg of OVA (grade V; Sigma-Aldrich, St Louis, MO, USA) in 100 µL of phosphate-buffered saline (PBS), or PBS alone, was applied to a sterile patch and attached to the tape-stripped area with an adhesive tape. Treatment was repeated once during the 1-week sensitization period, followed by a 2-week resting period before a repeat of the sensitization steps over 1 week. The mice were euthanized and samples were collected 24 hours after the last treatment (Supplementary Fig. S1).
Antibiotic treatment of the mice
Female BALB/c mice weighing 16-20 g (3 weeks old) were purchased from Orient Bio Inc. (Seongnam, Korea) and treated in accordance with the guidelines of the Institutional Animal Care and Use Committee of the Asan Medical Center and Ulsan University College of Medicine (identification code No. 2014-02-182).
For the induction of gut dysbiosis, mice were treated with an antibiotic cocktail in their drinking water for 2 weeks before primary sensitization with ampicillin (1 g/L; Ratiopharm, Ulm, Germany), vancomycin (500 mg/L; Cell Pharm, Hannover, Germany), ciprofloxacin (200 mg/L; Bayer Vital, Leverkusen, Germany), imipenem (250 mg/L; MSD, Haar, Germany), and metronidazole (1 g/L; Fresenius, Bad Homburg, Germany).25
Feces preparation and transplantation
To investigate the effect of fecal microbiota transplantation on gut dysbiosis, fecal samples were collected directly from untreated, age-matched, healthy donor mice. Fresh feces from 10 mice were pooled, mixed with sterile normal saline, and immediately homogenized. The homogenate was centrifuged at 100 × g for 5 minutes at 4°C and the supernatant was used for transplantation. The mice received 100 µL of the supernatant orally from 2 weeks before primary sensitization until the study endpoint.26
Probiotics preparation
The Lactobacillus rhamnosus (Lcr35) strain used in this study was obtained from Lyocentre® Laboratory (Aurillac, France) and prepared according to the manufacturer's directions. Lcr35 cells were suspended in saline and administered orally from 2 weeks before primary sensitization until the study endpoint.1126
Clinical scoring
Dorsum lesions were scored for erythema, scaling, and excoriation after each sensitization event using a 0-3 scoring system, where 0 = no lesion, 1 = mild lesion, 2 = moderate lesion, and 3 = severe lesion.11 The same 2 investigators randomly performed all scoring evaluations throughout the study.
Assessment of epidermal permeability barrier function
To determine whether epidermal permeability barrier function is altered as a result of OVA-induced AD, we measured transepidermal water loss (TEWL) at baseline at the beginning of the experiment, and then after each sensitization event, using a vapometer (SWL-3; Delfin Technologies Ltd, Kuopio, Finland).
Histology
The dorsal skin of the experimental mice was removed on the final day of the schedule, fixed in 10% phosphate-buffered formalin, and embedded in paraffin. Serial paraffin sections (4.5 mm thick) were stained with hematoxylin and eosin for the evaluation of edema.
Quantitation of immunoglobulin E (IgE) serum levels
Serum samples were obtained from blood taken during exsanguination of the mice after completing the sensitization, and stored at −80°C until use. Total IgE levels in sera were detected using the Mouse IgE ELISA kit (eBioscience, San Diego, CA, USA) in duplicate. The optical density was measured at 450 nm.
Real-time reverse transcriptase polymerase chain reaction (PCR)
To measure interleukin (IL) 4 expression in mouse skin, RNA was extracted from the dorsal skin of the experimental mice using the RNeasy kit (Qiagen, Valencia, CA, USA). Real-time PCR was performed using the TaqMan method on an ABI 7900 system (Applied Biosystems, Piscataway, NJ, USA). Each signal was normalized to that of GAPDH in the same sample.
Cell isolation and flow cytometry
Intraepithelial lymphocytes (IELs) were isolated from pooled mouse intestines as previously described.27 Briefly, the intestines were cut lengthwise into short segments and shaken in Roswell Park Memorial Institute (RPMI)-1640 containing 1 mM dithiothreitol, 2 mM ethylenediaminetetraacetic acid, and 2% (v/v) fetal calf serum (FCS) for 15 minutes at 37°C to remove the epithelial layer. The tissue remaining from the epithelial stripping was minced and digested in RPMI-1640 containing 1.5 mg/mL collagenase II (Gibco), 50 μg/mL DNase I (Sigma-Aldrich), and 1% FCS for 40-45 minutes at 37°C. The digested tissue was then washed and filtered at least twice to obtain a single-cell suspension. To harvest IELs from the epithelial layer, the cells were further spun through a 40:70 Percoll gradient, and IELs were isolated from the interphase layer. For intracellular cytokine staining, immediately after isolation, the cells were incubated for 4 hours with 50 ng/mL PMA (Sigma-Aldrich), 750 ng/mL ionomycin (Sigma-Aldrich), and 10 μg/mL GolgiPlug (BD Biosciences, Mountain View, CA, USA) in a tissue culture incubator at 37°C.
Next, the cells were fixed and permeabilized using the Intracellular Fixation & Permeabilization Buffer set from eBioscience and stained with the following antibodies: APC-anti-FOXP3 (FJK-16s), PE-anti-IL-17A (eBio17B7), and FITC-anti-CD4 (RM4-5) (all from eBioscience). For 3 innate lymphoid cell (ILC3) staining, isolated IELs were incubated with the following antibodies: APC-eFluor780-anti-CD19 (eBio1D3), PerCP-eFluor710-anti-CD3 (17A2), PE-anti-ROR gamma (t) (B2D), APC-anti-CD45 (30-F11), and FITC-anti-CD335 (NKp46) (29A1.4) (all from eBioscience). The stained cells were then analyzed by flow cytometry using a fluorescence-activated cell sorting (FACS) Calibur with CellQuest software (BD Biosciences).
Measurement of fecal SCFAs
For the measurement of SCFAs in mouse feces, 100 mg of fecal matter was first homogenized in 4 volumes of deionized water. Next, the supernatants were collected and an internal standard solution (100 μL of 10 mg/mL propionic acid-2,2-d2, Sigma-Aldrich Co. Ltd, Yongin, Korea), 20 μL of 0.6 N HCl, and 2 mL diethyl ether were added. The upper layer was then collected and the step was repeated as needed. After that, the solutions collected were completely dried under a vacuum and a 100-μL aliquot of 58 mg/mL pentafluorobenzyl bromide in acetonitrile and 20 μL of N,N-diisopropylethylamine were added to the derived matter. The mixtures were vortexed well and incubated at room temperature for 20 minutes. Finally, the sample solutions were vacuum-dried and reconstituted with 100 μL acetonitrile.
The SCFA levels were analyzed using a gas chromatography-mass spectrometry (GC-MS) system (7890A/5975A, Agilent) and HP-5 MS 30 m × 250 μm × 0.25 μm column (Agilent 19091S-433). Helium (99.999%) was used as the carrier gas. The initial temperature was 50°C and was raised to 100°C in 10°C/min increments after a 3-minute hold time. Next, the temperature was then raised to 130°C at 2.5°C/min and then to 230°C at 30°C/min which was maintained for 9 minutes. A 6-minute solvent delay and an SIM mode were applied. The extracted ion chromatogram corresponding to specific SCFAs was used for quantitation.
RESULTS
Oral antibiotic administration aggravates the clinical signs of AD in the mouse
A mouse model of AD was developed using repeated OVA sensitization and injury as previously described.24 An antibiotic cocktail was administered orally for 2 weeks to induce dysbiosis in the intestinal microbiota of the mice prior to OVA sensitization. Either fecal matter from healthy mice or probiotics (L. rhamnosus) was administered throughout the study to rescue the gut imbalance (Fig. 1).
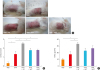 | Fig. 1Oral antibiotic administration aggravates clinical signs in a mouse model of AD. (A) Clinical features (erythema and swelling) in the AD mice treated as indicated. (B) TEWL levels. (C) Estimated clinical scores in AD mice treated as indicated.PBS, phosphate-buffered saline; OVA, ovalbumin; AT, oral antibiotic administration; TEWL, transepidermal water loss; AFT, antibiotics + orally administered fecal matter; APT, antibiotics + oral probiotics; AD, atopic dermatitis.
*P < 0.05, †P < 0.01, and ‡P < 0.001 by analysis of variance with Tukey's multiple comparison test.
|
Antibiotic treatment elicited significant swelling and erythematous skin lesions, as well as increased clinical scores, in the AD model mice (Fig. 1A and B). Significant TEWL was also observed in the antibiotic-treated AD mice (Fig. 1C). The OVA-sensitized groups treated with fecal matter from healthy mice and probiotics showed a significant improvement in clinical signs (Fig. 1A and B) and relevant suppressive pattern on TEWL (Fig. 1C).
Oral antibiotic administration up-regulates the systemic immune allergic response and Th2 cytokine levels in the skin
Antibiotic-treated AD mice showed a significantly increased total IgE level compared only to the OVA-sensitized animals and this effect was suppressed by treatment with both fecal matter from healthy mice and probiotics (Fig. 2A). The levels of the Th2 cytokine IL4 in the skin were assayed by real-time PCR and were found to be significantly increased in the antibiotic-treated AD mice compared to the control (Fig. 2B).
 | Fig. 2Oral antibiotic treatment increases the systemic immune allergic response and Th2 cytokine levels in the skin of AD mice. (A) Total IgE levels in the AD mice treated as indicated. (B) Measurement of IL4 expression in the skin of treated AD mice by real-time polymerase chain reaction.IgE, immunoglobulin E; PBS, phosphate-buffered saline; OVA, ovalbumin; AT, oral antibiotic administration; AFT, antibiotics + fecal administration orally; APT, antibiotics + oral probiotics; IL4, interleukin 4; AD, atopic dermatitis.
*P < 0.05, †P < 0.01, and ‡P < 0.001 by analysis of variance with Tukey's multiple comparison test.
|
Oral antibiotic administration suppresses the intestinal metabolites processed by the microbiota leading to reduced SCFA levels
To investigate whether antibiotics affect the intestinal metabolites processed by microbiota, SCFA levels in the guts of the AD mice were measured by GC-MS. Antibiotic treatment of OVA-sensitized animals led to a significant suppression of the SCFA levels, which re-covered with administration of fecal matter from healthy mice or probiotic treatment (Fig. 3). The effect of each treatment on the production of SCFAs was different. Notably, propionate (C3) and valerate (C5) were found to be significantly associated with the fecal and probiotic treatments in the AD mouse model (Fig. 3).
 | Fig. 3Oral antibiotic treatment suppresses the production of intestinal metabolites and SCFAs by microbiota. Gas chromatography-mass spectrometry SCFA measurements in atopic dermatitis mice treated as indicated.SCFA, short-chain fatty acid; PBS, phosphate-buffered saline; OVA, ovalbumin; AT, oral antibiotic administration; AFT, antibiotics + oral fecal administration; APT, antibiotics + oral probiotics.
*P < 0.05, †P < 0.01, and ‡P < 0.001 by analysis of variance with Tukey's multiple comparison test.
|
Oral antibiotic administration up-regulates inflammation associated with ILC3s and IL17+ cells and suppresses the levels of CD4+FOXP3+ cells in the gut
To investigate whether antibiotics affect the gut mucosal immune cells, we measured the levels of ILC3s and CD4+IL17+, and CD4+FOXP3+ regulatory T (Treg) cells in the small intestine of the AD mice by FACS. Antibiotics evoked a significant increase in the number of CD4+IL17+ cells (Fig. 4A and C) in the AD mice, but not administration of fecal matter from healthy mice or probiotic treatment (Fig. 4B and D). In addition, the CD4+FOXP3+ cell levels were significantly increased by treatment with fecal matter from healthy mice. The ILC3 number was also significantly increased by antibiotics and suppressed by fecal matter from healthy mice or probiotic treatment (Supplementary Fig. S2). These results suggested that the altered ILC3 and CD4+IL17+ cell profiles in the antibiotic-treated AD mice were representative of gut dysbiosis in these animals and that CD4+FOXP3+ cell levels can be used as a marker for the restoration of gut microbial homeostasis.
 | Fig. 4Oral antibiotic treatment increases IL-17+ cell-related inflammation and suppresses CD4+FOXP3+ cell levels in the gut of AD mice. (A, C) Population of CD4+IL-17+ cells in the small intestine of treated AD mice as determined by FACS. (B, D) Population of CD4+FOXP3+ cells in the small intestine of treated AD mice as determined by FACS.IL17, interleukin 17; PBS, phosphate-buffered saline; OVA, ovalbumin; AT, oral antibiotic administration; AFT, antibiotics + oral fecal administration; APT, antibiotics + oral probiotics; FACS, fluorescence-activated cell sorting; AD, atopic dermatitis.
*P < 0.05 and †P < 0.01 by analysis of variance with Tukey's multiple comparison test.
|
DISCUSSION
The results of our present study using an oral antibiotic cocktail-induced microbial depletion mouse model of AD indicate that intestinal microbial homeostasis may play a crucial role in the prevention of this condition. Following oral antibiotic treatment in our mouse model, AD phenotypes, including clinical score, TEWL, and IgE levels, were found to be aggravated and the expression level of the skin inflammatory cytokine IL4 was increased, whereas these parameters were improved by transplantation of feces from healthy mice or oral probiotic treatment. Our mechanistic results suggested that modulation of SCFA production by microbiota in the gut can regulate the CD4+IL17+ T cell/CD4+FOXP3+ Treg cell balance, thereby affecting the levels of ILC3 in the gut mucosa. Based on our current findings, the mechanisms underlying the development and treatment of AD should consider interactions between intestinal microbiota and other AD-related parameters.
In our AD mouse model, intestinal SCFA production was altered by fecal transplantation or oral probiotic treatment compared to that of antibiotic-treated animals. The levels of propionate and butyrate SCFAs showed significant increases when oral probiotics or healthy feces were administered to the mice. Notably, both propionate and butyrate have recently been reported to promote the accumulation of Treg cells in the colon by directly activating dendritic cells and T cells or G protein signals.2829
SCFAs are produced by gut microbiota as fermentation products and are required by intestinal epithelial cells to modulate immune responses to invading pathogens and commensal bacteria.10 Therefore, SCFAs prevent chronic intestinal inflammatory responses to microbes and their products. In our study, we found that the dysbiosis induced by antibiotic treatment elicits intestinal inflammation affecting the production of SCFAs in the intestine. The delicate balance between pro- (Th17) and anti-inflammatory (Treg) mechanisms essential for gut immune homeostasis is affected by the composition of the commensal microbial community and how they influence, for example, the SCFA profile.29
In the present study, however, we did not investigate the effects on the skin of oral antibiotic, fecal matter from healthy mice, or probiotic administration, and the effects of these treatments on AD require further investigation.
Intestinal bacteria play important roles in metabolism, immune cell development, epithelial cell integrity, motility, and prevention of gut colonization by pathogenic strains.30 We found that antibiotic treatments created an imbalance in the commensal intestinal microbiota, which could be a modern environmental factor with direct influence on intestinal homeostasis. Dysbiosis promotes the release by gram-negative bacteria of endotoxins and flagellin which bind to toll-like receptor 5 and induce intestinal damage and inflammation.3132 The proinflammatory cytokine IL17 has been reported to activate signaling pathways and induce the production of proinflammatory molecules that play major roles in neutrophil recruitment and tissue inflammation,33 particularly in autoimmune disease and intestinal cancer.3435 In our current analysis, the percentage of intestinal IL17+ T cells exhibited a significant increase in the antibiotic-treated mice, further indicating that antibiotics induce intestinal dysbiosis and IL17-related inflammation.
CD4+FOXP3+ Treg cells are known to maintain intestinal homeostasis34 and were significantly down-regulated by antibiotic treatment in the AD mice. The intestinal IL17+ T cell/FOXP3+ Treg cell imbalance was induced by alterations in the gut microbiota and was rescued by oral transplantation of healthy feces or administration of oral probiotics. Therefore, our present data indicate for the first time that an IL17+ T cell/CD4+FOXP3+ Treg cell imbalance in the gut could be a crucial determinant for AD development in early ages.
ILCs are the most recently described family of lymphoid cells and are known to regulate the development of lymphoid cells and intestinal inflammation,35 i.e., a larger number of ILC3s are associated with intestinal inflammation. These cells are also closely related to the regulation of mucosal adaptive immunity through Treg maintenance of intestinal homeostasis, supporting that ILC3s are also key in this process.3637
Both human and mouse ILC3s express IL22 and IL17, which play roles in tissue infection and inflammation responses during development.38
Our findings indicate that, like ILC3s, intestinal IL17+ T cell levels are significantly altered by antibiotic exposure; consequently, intestinal dysbiosis results in increased numbers of both cell types in mice. Butyrate has recently been reported to suppress the number of ILC3s in the intestine.36 Our current SCFA data revealed that probiotic treatment increases the percentage of butyrate and that, compared to antibiotics alone, co-administration of probiotics and antibiotics also reduced the ILC3 population levels in our mouse model. These observations demonstrate that oral probiotic administration promotes intestinal homeostasis by increasing the production of the SCFA butyrate, which modulates ILC3/Treg cell-related mechanisms.
Our current data on the significance of ILC3s are consistent with those from previous reports; however, due to the small number of available cells, further studies are needed to validate and confirm this relationship.
The present study has some limitations. It did not investigate the effects of antibiotics on skin changing microbiota and infections. However, the use of antibiotics (cocktail) was more massive in the study rather than in the treatment of pyoderma’ therefore, the effects could focus on eradication or strong strike of intestinal microbiota. The results in our AD model showed that the skin condition was controlled by systemic immune responses induced by changes in the intestinal microbiota rather than those in skin microbiome.
Skin commensal microbiota can also produce SCFAs and has similar immune regulatory mechanisms of the intestine by skin Tregs.39 The effect was not considered in the present study and focused on systemic immune modulation by dysbiosis of intestinal microbiota.
Additionally, the present study had not investigated direct changes in intestinal microbiome by effects of antibiotics and others. Instead, we investigated the SCFAs which are metabolites of microbiome. Based on the previous studies,81415 we showed the effects of antibiotics on dysbiosis of intestinal microbiota by SCFAs in mice. Further studies are needed to investigate changes in the intestine and skin microbiome resulting from antibiotic treatment in the AD model.
In conclusion, antibiotic use in early life needs to be considered in relation to the development or aggravation of AD mediated by gut microbial dysbiosis and associated changes in the SCFA profile. Guidelines are warranted to limit the use of antibiotics in patients at high risk of AD, the underlying mechanisms of which may be linked to IL17-positive and ILC3s.
 XML Download
XML Download