

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Breast cancer (BC) represents approximately 23% (1.38 million) of the total cancer cases and accounts for 14% (458,400) of cancer-related deaths [1]. The incidence of BC in developing countries throughout the Asia-Pacific regions is anticipated to continuously rise in the next few years [2]. Despite the advancement in the prognosis and treatment of BC, it is the most commonly detected cancer among women and the second leading cause of death in developed countries [3]. Comparison of cases among Western and Asian countries shows that BC is highly heterogeneous in terms of molecular, histological, and clinical features [4] as well as in terms of progression, pathology, and epidemiology [2]. Genomic alterations have been associated with the progression of BC, including genetic variations, genomic aberrations, and changes in splicing pattern, gene expression, and protein functionality. In recent years, efforts have been directed to investigate the pathogenesis of BC, as well as for its prevention and treatment. For instance, the St. Gallen Consensus Conference stated that neoadjuvant chemotherapy therapy was a preferred treatment approach for clinical stage II and III triple-negative breast cancer (TNBC) and human epidermal growth factor receptor 2 (HER2) positive BC [5]. Tono et al. [6] found that the combination of pertuzumab, trastuzumab, and eribulin mesylate could extend the overall survival of patients with HER2-positive metastatic BC. Furthermore, the study by Li et al. [7] showed that selective estrogen receptor modulators (SERMs) could reduce the risk of BC by affecting the expression of estrogen receptor (ER), as the persistent increase in the blood level of ER was found to increase the risk of BC. Moreover, it was reported that tumor-derived circulating extra vesicles (EVs) carrying breast cancer resistance protein (BCRP) contributed to the chemoresistance of BC [8].
Lysosome-associated protein transmembrane 4 beta (LAPTM4B) is a novel oncoprotein, which is upregulated in various human cancers, including breast, gastric, lung, esophageal, colon, rectal, and esophageal cancers and nonfunctioning pituitary adenoma [9]. LAPTM4B is reported to play important roles in many biological processes such as malignant transformation, apoptosis, invasion, and metastasis through its participation in multiple signal transduction pathways. The gene laptm4b is located on chromosome 8q22.1 with 7 exons and 6 introns [10], and has two alleles, laptm4b *1 and laptm4b *2. The laptm4b *1 allele has a single 19-bp segment, while laptm4b *2 allele has two tandem repeats of segments in close proximity [11].
The gene laptm4b is associated with cell proliferation and differentiation, and its overexpression causes uncontrolled cell growth and enhanced migration and invasion. Elevated LAPTM4B level was shown to contribute to the resistance to anthracyclines as well as recurrence [12], accelerated tumor progression, and disturbed the prognosis of BC [13]. Fan et al. found that laptm4b *2 was associated with the increased risk of BC among Chinese women [14]. LAPTM4B is also involved in the progression of other cancers and known to attenuate epirubicin-induced apoptosis of GBC-SD cells in vitro through a mitochondrion-dependent pathway [15]. LAPTM4B was reported as an independent factor in colorectal cancer prognosis [16], and laptm4b*2 allele was a cervical cancer risk factor that may increase the genetic susceptibility to cervical cancer in Chinese population [9]. In the present study, we determine the expression level of LAPTM4B in MCF-7 cell line and evaluate its influence on breast tumor cell proliferation, invasion, adhesion, and tube formation abilities in vitro and breast tumor progression in vivo.
METHODS
Cell lines and cell culture
The human BC cell line MCF-7 was cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS) (Invitrogen, Carlsbad, USA). All cell lines were maintained under standard cell culture conditions at 37°C in an incubator with 5% CO2 and 95% air.
Lentivirus-mediated gene transduction
Short-hairpin RNAs (shRNAs) targeting human laptm4b gene and its cDNA were inserted into a lentiviral vector (lenti-LAPTM4B-shRNA and lenti-LAPTM4B, respectively), which was co-transfected with helper plasmids into the human BC cell line MCF-7. Lentiviruses encoding targeted shRNA plasmids were produced by Neuron Biotech. The target sequence of LAPTM4B shRNA was TGCTGATCGGTAGCAGTTCCAAACACGTTTTGGCCACTGACTGACGTGTTTGGCTGCTACCGAT.
Western blot analysis
Cells were lysed with a lysis buffer (20 mM Tris-HCl, pH 7.5, 2 mM ethylenediaminetetraacetic acid, 1% NP-40, 150 mM sodium chloride [NaCl], 1 mg/mL sodium dodecyl sulfate, and 0.25 mg/mL sodium deoxycholate) supplemented with phenylmethylsulfonyl fluoride, phosphatase inhibitor, and protease inhibitor cocktail. Bicinchoninic acid protein assay kit (Pierce Biotechnology, Rockford, USA) was used to determine the protein concentration of each sample. Equivalent quantity of protein was separated with 12% sodium dodecyl sulfate polyacrylamide gels, and the separated bands were transferred onto nitrocellulose membranes. The membranes were blocked with 10% defatted milk and incubated overnight with an appropriate primary antibody. The membranes were washed and incubated with a corresponding horseradish peroxidase (HRP)-conjugated secondary antibody for 1 hour, followed by visualization with an enhanced chemiluminescence (ECL) system (Pierce Biotechnology, Rockford, USA). The primary antibodies used were as follows: anti-glyceraldehyde 3-phosphate dehydrogenase (Santa, Dallas, USA), anti-LAPTM4B (Abcam, Shanghai, China), anti-p21 (Santa), anti-cyclin-dependent kinase 1 (CDK1; Santa, 1:1,000), anti-vascular endothelial growth factor (Santa), anti-caspase-3 (CST, Danvers, USA), anti-matrix metalloproteinase (MMP)-2 (CST), anti-MMP-9 (CST), anti-P53 (CST), anti-phosphorylated (p)-protein kinase B (AKT) (Ser473) (Abcam), and anti-p-mammalian target of rapamycin (mTOR; Ser2448) (Abcam). The band grayscale of interested protein and loading control was quantified with ImageJ software (National Institutes of Health, Bethesda, USA), and the ratio of interested protein to loading control was calculated.
Cell proliferation assay
Cell Counting Kit (CCK)-8 assay was performed by culturing cells in 10% CCK-8 (Dojindo Laboratories, Kumamoto, Japan) reagent diluted in normal culture media at 37°C. Quantification was carried out using a microtiter plate reader (Thermo, Shanghai, China) after visual observation of the color change.
Cell cycle assay
Cell cycle and apoptosis were analyzed with flow cytometry, and the data were evaluated with FlowJo (FlowJo, LLC, Ashland, USA). The cells were centrifuged and the pellet obtained was washed once with 1× phosphate-buffered saline (PBS) + 0.1% bovine serum albumin (BSA). The supernatant was centrifuged again and discarded. The cell pellet obtained was treated with anti-mouse CD11c phycoerythrin (PE) (eBioscience, San Diego, USA) or PE anti-mouse CD206 (Biolegend, San Diego, USA), and the mixture was pipetted up and down several times to obtain a homogeneous cell mixture. The cells were incubated on ice for 30 min and centrifuged again. The resulted supernatant was discarded, and the cell pellet was washed twice with 1× PBS + 0.1% BSA. The cell pellet was diluted in cold 1× PBS, and the mixture was pipetted up and down to acquire a homogeneous cell mixture. The sample was immediately analyzed on an Epics Elite ESP (Coulter Electronics, Miami, USA) flow cytometer and the data were subsequently evaluated using the program Expo for Elite.
Transwell invasion assay
Matrigel invasion chambers (8 μm, 24-well cell culture inserts) were used with 5 × 104 cells resuspended in 500 μL DMEM. These cells were added to the upper chamber, while 500 μL DMEM with 10% FBS was added to the lower chamber. Cells on the upper surface of the membrane were removed after 22 hours and the membranes were stained with crystal violet. Cell numbers were determined by averaging cell counts from several separate fields.
Cell adhesion assay
Cells were treated with 10 mM ethylenediaminetetraacetic acid in DMEM, washed twice with DMEM, and resuspended in DMEM with 0.1% BSA at a concentration of 2 × 105 cells/mL. Each well of 96-well plates precoated with collagen I solution (1.6 μg/well in D-Hank's) was filled with a total of 100 μL cell suspension and incubated for 20 minutes at 37°C. Non-adherent cells were removed by washing four times with DMEM, followed by incubation for 4 hours in 100 μL DMEM with 10% FBS. Adherent cells were quantified with the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay.
In vivo tumorigenesis study
The animal experiment was carried out according to the guidelines of the Institution Animal Care and Use Committee, and all protocols were approved by East China University of Science and Technology. All applicable international, national, and/or institutional guidelines for the care and use of animals were followed. MCF-7 cells (2 × 106 cells in 1 mL PBS) were subcutaneously injected into the posterior flanks of 6-week-old female nude mice, and the tumor diameters were measured every 3 days. Tumor volume (mm3) was calculated as length × width2 × ½. Lentivirus was injected into the tumors at a volume of 50 μL every 3 days and a total of 5 times after the tumor size reached 50 mm3. Mice were euthanized after 1 week from the last injection, and tumors were excised and weighed.
Statistical analysis
Data were presented as mean ± standard deviation (SD) of separate experiments. The Student's t-test was used to determine significant differences between different groups, and all statistical analyses were performed with SPSS 16.0 (SPSS Inc., Chicago, USA). Statistical significance was defined as p < 0.05.
RESULTS
LAPTM4B promotes the proliferation of BC cells
To establish a model of human MCF-7 cells expressing different levels of LAPTM4B, we used a lentivirus carrying shRNA against human laptm4b or coding sequence (CDS) of human laptm4b to knockdown or overexpress laptm4b, respectively. The result of western blotting showed that LAPTM4B protein level decreased in the shRNA-treated group but increased in the overexpression group as compared with the non-transfected group (Figure 1A). These results also confirmed the specificity of LAPTM4B antibody used and the successful establishment of the cellular model.
Figure 1
LAPTM4B promotes the proliferation of breast tumor cells. (A) Relative levels of LAPTM4B in MCF-7 cells treated as indicated. Glyceraldehyde 3-phosphate dehydrogenase was used as the loading control. (B) Proliferation rates of MCF-7 cells treated as indicated. (C) Colony formation ability of MCF-7 cells treated as indicated. (D) Cycle distribution of MCF-7 cells treated as indicated.
LAPTM4B = lysosome-associated protein transmembrane 4 beta; NC = negative control; shRNA = short-hairpin RNA; OD = optical density.
*p < 0.05, †p < 0.01, ‡p <0.001.
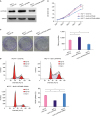
We obtained cellular growth curve and found a decrease in the growth rate for the cells lacking laptm4b expression. On the other hand, LAPTM4B overexpression increased the growth rate of cells as compared with the negative control groups (Figure 1B). This observation was consistent with the results of the colony formation assays that showed poor colony formation ability for LAPTM4B knockdown cells (Figure 1C). In addition, cell cycle analysis was carried out and LAPTM4B overexpression was found to significantly decrease the number of MCF-7 cells in G0/G1 phase, implying that LAPTM4B plausibly promoted the cells to enter into the S phase from the G1 phase (Figure 1D). These results demonstrate the positive regulatory effect of LAPTM4B on the proliferation and survival of breast tumor cells.
LAPTM4B promotes invasion and adhesion of breast tumor cells
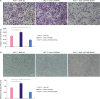
To measure the effect of LAPTM4B on the invasion and adhesion of MCF-7 cells, we conducted the transwell assays. In comparison with the laptm4b knockdown group, laptm4b overexpression group showed more number of cells invading from the upper chamber into the lower chamber (Figure 2A), which was in accordance with the result of the cell extracellular matrix adhesion assay. The laptm4b overexpression increased the number of adhesion cells (Figure 2B). From the transwell invasion and cell extracellular matrix adhesion assays, we conclude that laptm4b increased the invasion and adhesion capabilities of MCF-7 cells.
Figure 2
LAPTM4B promotes the invasion and adhesion of breast tumor cells. (A) Invasion ability of MCF-7 cells treated as indicated (scale bar = 100 μm); (B) Adhesion ability of MCF-7 cells treated as indicated (scale bar = 100 μm).
LAPTM4B = lysosome-associated protein transmembrane 4 beta; NC = negative control; shRNA = short-hairpin RNA; OD = optical density.
*p < 0.05, †p < 0.01, ‡p <0.001.
LAPTM4B promotes vasculogenic tube formation in breast tumor cells
To further clarify the role of laptm4b, we conducted the tube formation assay. Electron microscopy images clearly show that LAPTM4B-overexpressing MCF-7 cells had more branches (Figure 3A and B). This evidence further confirmed the positive effect of LAPTM4B on the invasion and adhesion of breast tumor cells. As vascular endothelial growth factor (VEGF)-A is a key player in the process of tumor angiogenesis, we compared the levels of secreted VEGF-A among three groups of MCF-7 cells expressing different levels of laptm4b. We found that laptm4b knockdown significantly decreased the level of secreted VEGF-A. On the contrary, the overexpression of this gene resulted in the upregulation in the level of VEGF-A. Thus, LAPTM4B inhibited the vasculogenic tube formation ability of tumors (Figure 3C).
Figure 3
LAPTM4B promotes vasculogenic tube formation in breast tumor cells. (A) Representative images of vasculogenic tubes formed in MCF-7 cells treated as indicated (scale bar = 100 μM). (B) Number of branch points formed in MCF-7 cells treated as indicated. (C) Concentration of VEGF secreted by MCF-7 cells treated as indicated. (D and E) Expression levels of various important regulators in MCF-7 cells treated as indicated.
LAPTM4B = lysosome-associated protein transmembrane 4 beta; NC = negative control; shRNA = short-hairpin RNA; VEGF = vascular endothelial growth factor; GAPDH = glyceraldehyde 3-phosphate dehydrogenase; Bcl = B cell lymphoma; CDK = cyclin-dependent kinase; HIF = hypoxia-inducible factor; MMP = matrix metalloproteinase; p = phosphorylated; AKT = protein kinase B; mTOR = mammalian target of rapamycin.
*p < 0.05, †p < 0.01.
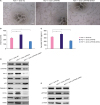
As the phosphoinositide 3-kinase (PI3K)/AKT/mTOR pathway is crucial for the regulation, growth, apoptosis, and motility of BC cells [1718], we investigated whether LAPTM4B had any effect on this pathway. We performed western blot analysis and found that the overexpression of LAPTM4B resulted in an increase in the protein levels of B cell lymphoma (Bcl)-2 (Figure 3D), p-AKT, and p-mTOR (Figure 3E). As MMPs are zinc-dependent endopeptidases participating in the process of angiogenesis, tumorigenesis, cancer invasion, and metastasis [1920], we also investigated whether LAPTM4B regulates MMP expression. Western blot data showed that both MMP2 and MMP9 levels were augmented in LAPTM4B-overexpressing MCF-7 cells, while the levels of both MMPs decreased in LAPTM4B knockdown MCF-7 cells (Figure 3D). Western blot data showed that LAPTM4B overexpression increased the expression of hypoxia-inducible factor (HIF)-1α, while the effect was opposite on LAPTM4B knockdown cells (Figure 3D). LAPTM4B knockdown also decreased the level of HIF-1α, MMP2, and MMP9, consistent with the finding that the knockdown of LAPTM4B expression inhibited the metastasis and angiogenesis of MCF-7 cells (Figure 3A and B). We explored the correlation between the protein levels of p21 and caspase-3 and LAPTM4B. Our data showed that p21 and caspase-3 levels were significantly reduced in MCF-7 cells overexpressing LAPTM4B, while opposite effects were observed in LAPTM4B knockdown cells (Figure 3D). These findings indicate that LAPTM4B promoted the proliferation of MCF-7 cells.
LAPTM4B promotes the progression of breast tumors in a nude mouse model
To elucidate the role of LAPTM4B in breast tumor progression, we conducted tumorigenicity assay in nude mice. MCF-7 cells stably expressing LAPTM4B shRNA or cDNA were separately injected into female mice to induce breast tumors. In comparison with the negative control groups, the group overexpressing LAPTM4B showed a significantly larger mean tumor volume and that injected with laptm4b knockdown cells showed a significantly smaller tumor volume (Figure 4A and B). The tumor samples were also collected to analyze the expression of protein markers such as p-AKT, p-mTOR, p21, and caspase-3 by western blot assay. In the laptm4b knockdown group, the expression of p-AKT and p-mTOR obviously decreased, while that of p21 and caspase-3 dramatically increased (Figure 4C). These results demonstrate that the decrease in the expression level of LAPTM4B could downregulate the signal transduction pathways related to tumor progression, suggesting that LAPTM4B promotes the progression of breast tumors in vivo.
Figure 4
LAPTM4B promotes the progression of breast tumors in vivo. (A) The size of tumors in different treatment groups. (B) Growth curves of tumors in different treatment groups. (C) Expression of various important proteins in tumors induced as indicated.
LAPTM4B = lysosome-associated protein transmembrane 4 beta; NC = negative control; shRNA = short-hairpin RNA; GAPDH = glyceraldehyde 3-phosphate dehydrogenase; p = phosphorylated; AKT = protein kinase B; mTOR = mammalian target of rapamycin; Bcl = B cell lymphoma; CDK = cyclin-dependent kinase; HIF = hypoxia-inducible factor.
*p <0.001.

DISCUSSION
Based on the current advanced technologies, many potential BC-related genes have already been investigated in detail. For instance, single-stranded conformational polymorphism (SSCP) was used to analyze mutations in the polymerase chain reaction (PCR) products of the DNA extracted from white blood cells. Researchers have identified the involvement of CYP1A1 and GSTP1 in breast carcinogenesis [21]. Baldwin et al. [22] developed a stable isotope labeling by amino acids in cell culture-based quantitative affinity purification/mass spectrometry approach to identify the interaction between PRMT1v1 and PRMT1v2 in BC cells.
Although the effect of LAPTM4B expression on the proliferation, migration, and invasion of BC cell lines has already been studied [23], its specific role in the progression of BC is still elusive. Here, we inspected the effects of different levels of LAPTM4B expression on the proliferation, invasion, adhesion, and tumor formation abilities of BC cells in vitro as well as on the breast tumor progression in vivo. Cell growth curves and colony formation assays showed that LAPTM4B promoted the proliferation of breast tumor cells, while cell cycle analysis revealed the underlying mechanism; LAPTM4B promoted the cells to enter from the G1 phase into the S phase. The transwell invasion and cell extracellular matrix adhesion assays showed that LAPTM4B overexpression increased the invasion and adhesion capabilities of MCF-7 cells. More branches were observed in LAPTM4B-overexpressing MCF-7 cells under an electron microscope. In comparison with LAPTM4B overexpression, LAPTM4B knockdown resulted in a decrease in the expression of VEGF-A and significantly inhibited vasculogenic tube formation in tumors. These results were also verified with western blot analysis. Thus, we conclude that LAPTM4B influenced the regulation, growth, apoptosis, and motility of MCF-7 cells via the AKT/mTOR pathway. The knockdown of LAPTM4B expression resulted in a decrease in the level of p-AKT and p-mTOR, thereby affecting the dissociation and decreasing the expression of Bcl-2. The possible explanation was that activated AKT could phosphorylate BAD that formed a protein complex with Bcl-2, thereby allowing the binding of BAD to 14-3-3 protein and consequently releasing Bcl-2 [24]. MMP2 and MMP9 are involved in angiogenesis, tumorigenesis, cancer invasion, and metastasis, and HIF-1α could accelerate mammary tumor progression and metastasis. Overexpression of LAPTM4B in MCF-7 cells significantly upregulated the expression levels of HIF-1α, MMP2, and MMP9. Nude mice transplanted with LAPTM4B-overexpressing MCF-7 cells produced larger tumors and showed higher levels of HIF-1α than the control mice. Thus, LAPTM4B promoted invasion, adhesion, and angiogenesis by stimulating the expression of HIF-1α, MMP2, and MMP9. p21 (WAF1/CIP1) is a tumor suppressor that participate in the regulation of cell cycle and cell proliferation, while caspase-3 seems to be a central mediator of programmed cell death in mammalian cells. Both these proteins were significantly downregulated in MCF-7 cells overexpressing LAPTM4B, but their levels dramatically increased in LAPTM4B knockdown cells.
Further experiments are warranted to determine the downstream effectors and upstream regulators of the AKT/mTOR pathway. Whether LAPTM4B activates AKT/PI3K could be verified using a PI3K inhibitor [25]. Fangchinoline was found to decrease the phosphorylation of AKT and glycogen synthase kinase-3β and increase the level of active caspase-3 in MDA-MB-231 BC cells [26]. Tang et al. [27] showed that ursolic acid downregulated the expression of survivin by blocking PI3K/AKT and activating caspase-3. These studies offer the plausible explanation on how the AKT/mTOR pathway affects the expression of caspase-3 in MCF-7 cells. Further investigation is needed to address whether the differential expression of MMP2 and MMP9 caused by LAPTM4B was related to p21-activated kinases [28]. The possible downstream effectors and the upstream regulators of AKT/mTOR pathway could also be identified by mass spectrometry via screening of differentially expressed proteins.
In summary, we found that LAPTM4B regulated the components of the AKT/mTOR pathway and promoted the proliferation of MCF-7 cells by decreasing the expression of p21 (WAF1/CIP1) and caspase-3. In addition, it promoted the invasion, adhesion, and angiogenesis of BC cells by stimulating the expression of HIF-1α, MMP2, and MMP9. LAPTM4B may thus serve as a potential therapeutic target for BC treatment.
 XML Download
XML Download