

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Breast cancer is the most frequently diagnosed cancer and is the second leading cause of cancer mortality in women worldwide after lung cancer. Breast cancer commonly exhibits invasive malignancy, shows increased incidence with age, and poses a serious global health problem [1]. Most breast cancer cells originate from the milk-forming duct of the mammary gland in epithelial tissues. The cancer cannot be completely eradicated because of its high degree of genetic heterogeneity and mutations, although the patient survival rate has increased in recent decades due to medical advances [2]. Based on gene expression profiling patterns, breast cancer can be classified into 4 main molecular subtypes, namely, luminal A, luminal B, human epidermal growth factor receptor 2 (HER2)-positive, and basal-like [3]. Specifically, the basal-like subtype is classified as triple-negative breast cancer (TNBC) because it is negative for the estrogen receptor (ER), progesterone receptor, and HER2. TNBC is associated with a poorer prognosis, a higher rate of distant recurrence, and the shortest overall survival rate of all breast cancers, and is also less sensitive to typical endocrine therapies than other subtypes owing to its aggressiveness and the limited therapeutic options [4].
Numerous therapeutic approaches have been developed for cancer treatment. Typical standard therapy includes radiotherapy, chemotherapy, and/or radiochemotherapy followed by surgery. Several molecular targeting agents have been developed and used clinically in combination with radiotherapy to increase therapeutic efficacy [56]. Tamoxifen and trastuzumab are the best-known chemotherapeutic drugs for targeted treatment of breast cancer. Although these agents improve clinical outcomes in patients with ER-positive and metastatic HER2-positive breast cancer, they may also elicit adverse effects and resistance to therapy [78], highlighting the importance of developing novel targeted drugs. Various types of cancer, including breast cancer, often resulted from disrupted regulation of complex signaling networks. Extensive surveys of primary tumors and tumor-derived cell lines have demonstrated that Janus kinase (JAK)/signal transducer and activator of transcription 3 (STAT3) signaling is constitutively activated in various cancer types and is recognized as a valuable therapeutic target for cancer treatment. In particular, JAK2/STAT3 signaling is regularly observed in a variety of tumor-derived cancer cells and cancer patients [9]. The transcription factor STAT3 is frequently related to cancer aggressiveness and malignancy; its activation in the inflammation-associated tumor microenvironment promotes cancer cell survival, proliferation, invasion, and metastasis, as well as the maintenance of cancer stem cell (CSC) traits in a wide range of cancers [10].
Tubulosine is first isolated from the bark of Pogonopus tubulosus in 1964 [11]. Although the structure of tubulosine and how to isolate it have long been known, its biological and pharmacological functions have yet to be elucidated. In this study, we demonstrated the potent anticancer activity of tubulosine, as well as the associated molecular mechanisms in breast cancer cells.
METHODS
Reagents
The chemical library was obtained from the Developmental Therapeutics Program (DTP) of the National Cancer Institute (NCI), where it is available to investigators for non-clinical research purposes. Information on compound synthesis and purity is not available from the DTP/NCI website. Interleukin (IL)-6 and epidermal growth factor (EGF) were obtained from PeproTech (Rocky Hill, USA). Cholera toxin, hydrocortisone, insulin, and the pan-JAK inhibitor AG-490 were purchased from Sigma-Aldrich (St. Louis, USA). All other chemicals used were of analytical grade and were obtained from Sigma-Aldrich unless otherwise noted.
Cell lines
The human mammary breast epithelial cell line MCF-10A was obtained from the American Type Culture Collection (Manassas, USA) and maintained in Dulbecco's Modified Eagle Medium (DMEM)/F-12 medium supplemented with 5% horse serum, 100 ng/mL cholera toxin, 20 ng/mL EGF, 0.5 mg/mL hydrocortisone, 10 µg/mL insulin, and 1% penicillin/streptomycin (HyClone, Pittsburgh, USA). The human breast cancer cell lines Hs578T, MCF-7, MDA-MB-231, and MDA-MB-468 were obtained from the American Type Culture Collection and maintained in DMEM, Eagle's Minimal Essential Medium, or Leibovitz's L-15 medium supplemented with 10% fetal bovine serum (FBS) and 1% penicillin/streptomycin. All the human cell lines were maintained at 37°C in a humidified incubator containing 5% CO2. Parental macrophage-like Drosophila Schneider S2-NP cells were maintained in Schneider's Drosophila medium (Gibco BRL, Grand Island, USA) supplemented with 10% FBS and 1% penicillin/streptomycin in an incubator at 25°C. S2-NP-STAT protein at 92E (STAT92E) cells stably expressing both the p10 × STAT92E-firefly luciferase and promoter-associated RNA polymerase III-Renilla luciferase reporter constructs [12] were maintained in the same medium supplemented with 500 μg/mL geneticin (Sigma-Aldrich).
Cell-based STAT92E reporter assay
S2-NP-STAT92E cells were co-cultured with Unpaired (Upd)-producing S2-NP cells prepared through transient transfection with an actin promoter-driven upd expression plasmid using Effectene transfection reagent (Qiagen, Valencia, USA). The cells were incubated for 24 hours in the presence of the vehicle alone or 10 µM of each test compound, as previously described [12]. The STAT92E-luciferase reporter activity was determined by measuring relative luciferase luminescence units using the Dual-Glo Luciferase Assay System (Promega, Madison, USA); firefly luciferase activity was normalized to that of Renilla luciferase.
Cell viability assay
S2-NP-STAT92E cells were co-cultured with Upd-producing S2-NP cells in a 96-well plate for 24 hours in the presence of the vehicle alone or various concentrations of tubulosine. Ten microliters of EZ-CyTox Enhanced Cell Viability Assay Reagent (Daeil Lab Service, Seoul, Korea) was added and soluble formazan was measured based on absorbance at 450 nm using a microplate reader (Molecular Devices, Sunnyvale, USA) following incubation for 2–4 hours at 37°C. Breast cancer cells were seeded at a density of 1 × 104 cells per well in a 96-well plate and incubated in culture medium until 70%–80% confluence. The cells were treated with tubulosine in the presence or absence of IL-6 for 24–72 hours, and cell viability was then determined from the absorbance at 450 nm after adding EZ-CyTox Enhanced Cell Viability Assay Reagent.
Western blot analysis
Protein samples were separated using sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS–PAGE) and transferred onto nitrocellulose membranes (Pall Corp., Port Washington, USA). The membranes were blocked for 1 hours in blocking buffer containing 5% skimmed milk in Tris-buffered saline containing 0.1% Tween 20 and subsequently incubated with the corresponding primary antibodies overnight at 4°C. Blots were washed and incubated with horseradish peroxidase-conjugated secondary antibodies at room temperature for 2 hours, following which the signals were detected using ECL reagent (SurModics, Eden Prairie, USA). Antibodies specific for phospho-JAK1, JAK1, phospho-JAK2, JAK2, phospho-JAK3, JAK3, phospho-tyrosine kinase 2 (TYK2), TYK2, phospho-STAT1, STAT1, phospho-STAT3, poly (ADP-ribose) polymerase (PARP), cleaved caspase 3, B-cell lymphoma (Bcl)-2, Bcl-xL, survivin, and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) were obtained from Cell Signaling Technology (Danvers, USA). Antibodies specific for STAT3, IL-6 receptor α (IL-6Rα), and glycoprotein 130 (gp130) were obtained from Santa Cruz Biotechnology (Santa Cruz, USA), while those for PY-20 and HA were obtained from Abcam (Cambridge, USA).
Immunoprecipitation assay
S2-NP cells were transiently transfected with an HA-tagged STAT92E expression plasmid. The cells were co-cultured with Upd-producing S2-NP cells in the presence of the vehicle alone or tubulosine for 24 hours. Whole-cell lysates were incubated with the anti-HA antibody at 4°C overnight and then immunoprecipitated with protein G-sepharose (GE Healthcare, Pittsburgh, USA) at 4°C for 2 hours. The immune complexes were subjected to SDS–PAGE and probed with anti-PY-20 and anti-HA antibodies to detect the levels of STAT92E and phospho-tyrosine STAT92E. In addition, MCF-7 cells were incubated for 3 hours with the vehicle alone or tubulosine and then stimulated with IL-6 for 10 minutes. Immunoprecipitation was performed with whole-cell lysates using anti-IL-6Rα or anti-gp130 antibodies, followed by Western blot analyses.
Semi-quantitative reverse transcription polymerase chain reaction (RT-PCR)
Total RNA was isolated using TRIzol Reagent (Invitrogen, Carlsbad, USA) and semi-quantitative RT-PCR was performed using AccuPower RT-PCR PreMix (Bioneer, Daejeon Korea) according to the manufacturer's protocol. Briefly, total RNA was reverse transcribed using a thermal cycler programmed at 42°C for 1 hour and PCR was performed with 30 cycles at 94°C for 30 seconds, 58°C for 30 seconds, and 72°C for 30 seconds, and a final extension step at 72°C for 5 minutes. The amplicons were resolved on agarose gels with DNA SafeStain (Lambda Biotech, St. Louis, USA) and visualized using a Gel Doc 2000 imaging system (Bio-Rad Laboratories, Hercules, USA). The primers used in this experiment were matrix metalloproteinase (MMP)-3 (QT00060025), MMP-9 (QT00040040), MMP-11 (QT00024031), Bcl-2 (QT00025011), Bcl-xL (QT00236712), SURVIVIN (QT01679664), and GAPDH (QT0007924) and were all obtained from Qiagen.
Immunofluorescence staining
Cells cultured in lysine-coated dishes were incubated for 6 hours in the presence of the vehicle alone or tubulosine and then stimulated with IL-6 for 30 minutes. The cells were permeabilized with 0.1% Triton X-100 in PBS for 15 minutes, fixed with 4% paraformaldehyde for 15 minutes, and then blocked with 0.1% Triton X-100 containing 3% bovine serum albumin for 1 hours. The dishes were incubated at 4°C overnight with anti-phospho-tyrosine STAT3 (1:100) and anti-STAT3 (1:100) antibodies. After washing with PBS, the dishes were incubated with the appropriate secondary antibodies at room temperature for 2 hours. Nuclei were counterstained with 4′,6-diamidino-2-phenylindole (DAPI) and images were acquired using a Zeiss Axiovert 200 inverted fluorescence microscope with the LSM 510 META confocal microscope system (Carl Zeiss, Oberkochen, Germany).
Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay
Cells were seeded at a density of 5 × 104 cells per well in a 12-well plate and then incubated for 48 hours with the vehicle alone or tubulosine in the presence or absence of IL-6. To perform the TUNEL assay, cells were stained using an In Situ Apoptosis Detection kit (Clontech, Mountain View, USA) according to the manufacturer's protocol, and fluorescein isothiocyanate signals were visualized using confocal laser scanning microscopy (Carl Zeiss).
Statistics
All statistical analyses were performed with the 2-tailed Student's t-test, which was conducted using the 2-way ANOVA function in GraphPad Prism 5.0 (GraphPad Software, San Diego, USA). Each experiment was performed independently at least in triplicate and the data are represented as means ± standard deviation. Values of p < 0.05 were considered statistically significant.
RESULTS
Identification of tubulosine as an inhibitor of STAT92E activity in Drosophila cells
We previously reported that the cultured Drosophila cell line S2-NP is a useful tool for identifying small-molecule inhibitors of JAK/STAT signaling [12], as the Drosophila JAK/STAT signaling pathway consists of only a single core component with the same cellular functions as in mammals [13]. To identify small-molecule inhibitors of JAK/STAT signaling, we performed cell-based high-throughput screening using S2-NP-STAT92E cells and a diverse set of NCI compounds.
Tubulosine (Figure 1A) was the strongest inhibitor of STAT92E transcriptional activity among the tested compounds. Tubulosine strongly suppressed the Upd-induced transcriptional activity of STAT92E in a concentration-dependent manner, with an IC50 value of 54.5 nM (Figure 1B). We further examined whether tubulosine could inhibit the tyrosine phosphorylation of STAT92E, which corresponds to the activity of the STAT protein in mammals. The level of STAT92E tyrosine phosphorylation was determined using an anti-PY-20 antibody, followed by immunoprecipitation with an anti-HA antibody. Tubulosine treatment resulted in a concentration-dependent decrease in STAT92E phosphorylation in Upd-induced S2-NP cells (Figure 1C). Moreover, exposure to tubulosine also led to a concentration-dependent reduction in the viability of Upd-induced S2-NP cells (Figure 1D). These results indicate that tubulosine is a small-molecule inhibitor of STAT92E signaling in Drosophila cells.
 | Figure 1Tubulosine inhibits Drosophila STAT92E activity. (A) The chemical structure of tubulosine. (B) Tubulosine inhibits Upd-induced STAT92E transcriptional activity. S2-NP-STAT92E cells were co-cultured with Upd-producing S2-NP cells for 24 hours in the presence of the vehicle (0.1% dimethyl sulfoxide) alone or the indicated concentrations of tubulosine. STAT92E reporter activity was normalized using the ratio of firefly luciferase to Renilla luciferase activity. Reporter activity without Upd stimulation was set to 1. (C) Tubulosine inhibits Upd-induced STAT92E tyrosine phosphorylation. S2-NP cells were incubated with various concentrations of tubulosine for 24 hours in the presence or absence of Upd. Immunoprecipitation and Western blot analyses were performed to determine the levels of pY-STAT92E and STAT92E. (D) Cells were prepared as described in (B), and cell viability was determined using EZ-CyTox Enhanced Cell Viability Assay reagent. Cell viability was represented as a control % compared to the vehicle-treated group. Results are represented as means ± standard deviation of 3 independent experiments (n = 3).STAT92E = signal transducer and activator of transcription protein at 92E; Upd = Unpaired.
*p < 0.005 compared to the vehicle-treated group; †p < 0.05 and ‡p < 0.005 compared to the Upd-induced group.
|
Tubulosine inhibits IL-6-induced JAK2/STAT3 signaling
We further examined whether tubulosine could inhibit JAK/STAT3 signaling in human breast cancer cell lines. Among the cell lines tested, the TNBC-subtype cell lines like MDA-MB-231 and MDA-MB-468 expressed persistently active forms of STAT3, JAK1, and JAK2 compared to the mammary breast epithelial MCF-10A cell line and the ER-positive Hs578T and MCF-7 breast cancer cell lines (Supplementary Figure 1). These results suggest that highly activated JAK1/JAK2/STAT3 signaling plays an important role in maintaining breast cancer malignancy and reduced sensitization to cancer therapy, and that this signaling pathway may be a critical therapeutic target.
To determine the effects of tubulosine on JAK/STAT3 signaling, various breast cancer cell lines were incubated with tubulosine and then stimulated with IL-6. The IL-6 protein is a major proinflammatory cytokine produced mainly by immune cells, cancer cells, and surrounding tumor-associated cells, like fibroblasts, in the inflammation-associated tumor microenvironment. IL-6 plays an essential role in cancer progression and tumor malignancy by activating JAK2/STAT3 signaling [14]. Stimulation of IL-6 led to a marked increase in the level of tyrosine-phosphorylated JAK2 and STAT3 compared to non-stimulated parental cells. In parallel with Upd-associated induction of Drosophila S2-NP cells, tubulosine treatment resulted in a reduction of the elevated levels of STAT3 and JAK2 tyrosine phosphorylation in IL-6 stimulated MCF-10A, Hs578T, and MCF-7 cells (Figure 2A), as well as in IL-6-stimulated MDA-MB-231 and MDA-MB-468 cells (Figure 2B). Moreover, exposure to tubulosine also resulted in a concentration-dependent reduction in STAT3 tyrosine phosphorylation levels in IL-6-stimulated MCF-10A and MCF-7 cells (Figure 2C).
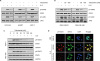 | Figure 2Tubulosine inhibits IL-6-induced JAK2/STAT3 signaling in breast cancer cells. (A-C) Cells were incubated for 6 hours in the presence of the vehicle (0.1% dimethyl sulfoxide) alone or tubulosine (100 nM or the indicated concentrations) and then stimulated with IL-6 (20 ng/mL) for 10 minutes. Protein samples were prepared from whole-cell lysates and Western blot analyses were performed. GAPDH was used as a loading control. (D) Cells were prepared as described in Figure 2A-C, followed by IL-6 stimulation for 30 minutes and then performed immunofluorescence staining. Nuclei were counterstained with DAPI.IL = interleukin; JAK2 = Janus kinase 2; STAT3 = signal transducer and activator of transcription 3; PBS = phosphate-buffered saline; GAPDH = glyceraldehyde 3-phosphate dehydrogenase; DAPI = 4′,6-diamidino-2-phenylindole.
|
Tyrosine-phosphorylated STAT3 can homodimerize and translocate to the nucleus, where it transcriptionally induces numerous target genes by binding to specific DNA sequences [10]. Therefore, we further examined the nuclear translocation of tyrosine-phosphorylated STAT3 using immunofluorescence staining. Consistent with the results of STAT3 tyrosine phosphorylation, IL-6 stimulation led to a marked increase in the levels of tyrosine phosphorylation and nuclear translocation of STAT3 in MCF-7 cells, whereas tubulosine treatment elicited a marked reduction in these levels (Figure 2D). These results clearly indicate that tubulosine is a potent inhibitor of JAK2/STAT3 signaling induced by stimulation with the proinflammatory cytokine IL-6, which is produced in the inflammation-associated tumor microenvironment.
Tubulosine blocks the binding of IL-6Rα and gp130
The above results led us to further investigate the mechanism through which tubulosine inhibits IL-6-induced JAK2/STAT3 signaling. IL-6-induced signaling cascades are mediated by the IL-6-specific receptor IL-6R and gp130. The IL-6/IL-6R/gp130 complex is formed following the binding of IL-6 to IL-6R [15] and targeting the formation of this complex is a potential therapeutic approach for breast cancer treatment [16]. Before examining the mechanism of tubulosine activity, we first assessed the time point at which tubulosine inhibits IL-6-stimulated STAT3 tyrosine phosphorylation. Tubulosine treatment resulted in a time-dependent decrease in STAT3 phosphorylation, and effective inhibition was achieved within 2 hours (Figure 3A). Next, we performed immunoprecipitation assays using anti-IL-6Rα and anti-gp130 antibodies. IL-6 stimulation increased the binding of IL-6Rα and gp130, but this binding was effectively suppressed in the presence of tubulosine (Figure 3B and C). These results indicate that the inhibition of IL-6-induced JAK2/STAT3 signaling by tubulosine is mediated by the blocking of IL-6Rα and gp130 binding.
 | Figure 3Tubulosine blocks the binding of IL-6Rα and gp130. (A) MCF-7 cells were incubated with tubulosine (100 nM) for the indicated periods and then stimulated with IL-6 (20 ng/mL) for 10 minutes. Protein samples were prepared from whole-cell lysates and Western blot analyses were performed. GAPDH was used as a loading control. (B and C) MCF-7 cells were incubated for 3 hours with the vehicle (0.1% dimethyl sulfoxide) alone or tubulosine and then stimulated with IL-6 (20 ng/mL) for 10 minutes. Immunoprecipitation was performed for whole-cell lysates using an anti-IL-6Rα (B) or anti-gp130 antibody (C) followed by Western blot analyses.STAT3 = signal transducer and activator of transcription 3; IL = interleukin; IL-6Rα = interleukin-6 receptor α; GAPDH = glyceraldehyde 3-phosphate dehydrogenase.
|
Tubulosine induces apoptotic cell death
Hyperactivated STAT3 signaling is associated with increased survival and proliferation of cancer cells, suggesting that tubulosine can decrease the viability of IL-6-induced breast cancer cells. To address this possibility, we first performed cell viability assays on MCF-7, MDA-MB-231, and MDA-MB-468 cells, which were incubated for up to 72 hours with tubulosine in the presence of IL-6. Cell viability decreased significantly with tubulosine treatment in a time-dependent manner in all tested cell lines (Figure 4A). Similarly, the pan-JAK/STAT3 signaling inhibitor AG-490 [17] also decreased cancer cell viability in a time-dependent manner. We further performed a TUNEL assay to determine whether this reduced cell viability was associated with apoptotic cell death. MCF-7 cells were incubated for 48 hours with the vehicle alone or tubulosine in the presence or absence of IL-6. Tubulosine treatment resulted in a larger apoptotic cell population among IL-6-stimulated cells compared to vehicle-treated control cells (Figure 4B).
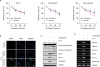 | Figure 4Tubulosine induces apoptotic cell death. (A) Cells were incubated with tubulosine (100 nM) or AG-490 (150 µM) for the indicated periods in the presence of IL-6 (20 ng/mL). Cell viability was determined using EZ-CyTox Enhanced Cell Viability Assay reagent and represented as a control % compared to the vehicle-treated group. Results are represented as means ± SD of three independent experiments (n = 3). (B and C) MCF-7 cells were incubated for 48 hours with the vehicle (0.1% DMSO) alone or tubulosine (100 nM) in the presence or absence of IL-6 (20 ng/mL). A TUNEL assay (B) and Western blot analyses (C) were performed. (D) MCF-7 cells were incubated for 3 hours with the vehicle (0.1% DMSO) alone or tubulosine (100 nM), and further incubated for 12 hours in the presence or absence of IL-6 (20 ng/mL). Semi-quantitative RT-polymerase chain reaction was performed using target-specific primers.TUNEL = terminal deoxynucleotidyl transferase dUTP nick end labeling; DAPI = 4′,6-diamidino-2-phenylindole; DMSO = dimethyl sulfoxide; IL = interleukin; Bcl = B-cell lymphoma; PARP = poly (ADP-ribose) polymerase; MMP = matrix metalloproteinase.
*p < 0.05 and †p < 0.005 compared to the vehicle-treated group.
|
To further clarify the molecular mechanisms through which apoptotic cell death in tubulosine-treated cancer cells increased, we performed Western blotting and semi-quantitative RT-PCR analyses. Tubulosine treatment combined with IL-6 stimulation resulted in increased fragmentation of PARP and caspase 3 (Figure 4C), which are hallmark proteins of apoptosis [18]. In addition, there was also a reduction in the protein and mRNA levels of antiapoptotic proteins like Bcl-2, Bcl-xL, and survivin (Figure 4C and D). Interestingly, we also observed reduced mRNA levels of MMPs, including MMP-3, MMP-9, and MMP-11 (Figure 4D), that are potential biomarkers of invasion and metastasis in breast cancer through degradation of the extracellular matrix [19]. These results indicate that inhibition of IL-6-induced JAK2/STAT3 signaling by tubulosine decreases cancer cell survival by inducing apoptosis, and also decreases invasion and metastasis by cancer cells in the inflammation-associated tumor microenvironment.
DISCUSSION
Identifying pharmacologically active compounds targeting specific signaling pathways is an important step in early-stage drug discovery. In this study, we performed cell-based high-throughput screening to identify JAK/STAT signaling inhibitors using cultured Drosophila cells. We identified tubulosine as a potent JAK2/STAT3 signaling inhibitor, and showed that it blocked the binding of the IL-6 receptors IL-6Rα and gp130, thereby inhibiting JAK2/STAT3 signaling in IL-6-induced breast cancer cells. Additionally, tubulosine induced apoptotic cell death and decreased invasion and metastasis by cancer cells (Figure 5).
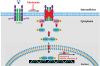 | Figure 5Schematic diagram representing the proposed mechanism of action of tubulosine. Tubulosine inhibits IL-6-induced JAK2/STAT3 signaling by blocking IL-6Rα and gp130 binding.IL = interleukin; IL-6Rα = interleukin-6 receptor α; JAK2 = Janus kinase 2; STAT3 = signal transducer and activator of transcription 3; gp130 = glycoprotein 130.
|
The JAK/STAT signaling cascade and the cellular functions of the core components of each step are highly evolutionarily conserved, from invertebrates to vertebrates [13]. Importantly, the JAK/STAT signaling pathway of the fruit fly Drosophila melanogaster consists of 3 highly related Upd ligands (Upd1, 2, and 3), 1 receptor (Domeless), 1 JAK (Hopscotch), 1 STAT (STAT92E), and 2 negative regulators of each, namely, suppressor of cytokine signaling at 36E and protein inhibitors of activated STAT [13]. Consequently, Drosophila is often used as a simple model organism for evaluating signaling pathways and identifying pharmacologically active small molecules. For cell-based high-throughput screening to identify small molecule inhibitors targeting JAK/STAT signaling, we used S2-NP-STAT92E cells constructed from the parental macrophage-like Drosophila S2-NP cells through transfection with both the 10×STAT92E-firefly luciferase and the RNA polymerase III-Renilla luciferase reporter genes [12].
Breast cancer is the most frequently diagnosed cancer in women worldwide. It is a serious public health concern and its treatment poses numerous challenges. Breast cancer is divided into 4 stages, classified based on tumor size, the number of metastasis-positive lymph node stations, and the degree of cancer invasion into other areas of the body [20]. In addition, there are 4 subtypes of breast cancer, namely: luminal A, luminal B, HER2-positive, and basal-like, that are classified based on gene expression patterns [3]. Treatment for breast cancer is commonly determined by the stage and subtype, and may involve radiation, chemotherapy, or a combination of both following surgery, indicating that numerous therapeutic options are needed. Based on analyses of JAK/STAT signaling, persistently active JAK1/JAK2/STAT3 signaling was observed in the TNBC cell line subtype. This signaling may be associated with reduced efficacy of TNBC therapy as patients with this subtype have shorter survival rates, higher recurrence rates, comparatively highly aggressive cancer, and reduced sensitivity to treatment [4].
Numerous proinflammatory cytokines are produced by immune cells, cancer cells, and cancer-associated stromal cells, such as fibroblasts, in the inflammation-associated tumor microenvironment [10]. IL-6 is a representative proinflammatory cytokine that promotes invasiveness, tumor malignancy, drug resistance, and CSC properties by activating IL-6R/gp130-mediated JAK2/STAT3 signaling [1421]. IL-6 is expressed in approximately 50% of breast cancers and its level is higher in TNBC cell lines than in breast cancer cells of other subtypes. In addition, the growth of TNBC cells depends on autocrine IL-6 signaling [22]. Together, this evidence indicates that targeting IL-6-induced JAK2/STAT3 signaling is an important therapeutic option for the treatment of TNBC. IL-6 binds to 2 different types of receptor molecules, IL-6R and gp130 [15]. The IL-6-induced signaling pathway is mediated by the formation of a hexameric IL-6/IL-6R/gp130 complex composed of 2 subunits of each molecule [23].
Numerous inhibitors have been developed to directly block IL-6/IL-6R/gp130 complex formation in cancer therapy [16]. Among them, LMT-28 was recently developed as a direct inhibitor targeting the binding of gp130 to IL-6/IL-6Rα [24]. Here, we newly identified tubulosine as a direct inhibitor that blocks IL-6Rα and gp130 binding in IL-6-induced breast cancer cells. Tubulosine exposure resulted in inhibition of the JAK2/STAT3 signaling pathway, thereby inducing apoptosis, decreasing viability, and reducing invasion and metastasis by cancer cells.
Tubulosine is first isolated from the bark of P. tubulosus in 1964 [11]. The detailed chemical properties of tubulosine and its antitumor activity were further evaluated following isolation from the sap of Pogonopus speciosus in the 1990s [2526]. Its antitumor activity has also been evaluated by measuring its cytotoxicity against various cancer cell lines, including BC1, Lu1, Col2, KB, KB-V+, KB-V−, LNCaP, SW626, SKNSH, and M109. Additional biological and pharmacological functions of tubulosine include amebicidal activity and inhibition of protein biosynthesis [27], inhibition of eukaryotic elongation factor 2-dependent peptide chain elongation [28], and inhibition of hypoxia-inducible factor-1A transcriptional activity [29]. Here, we demonstrated a novel mechanism of action of the anticancer activity of tubulosine, which is associated with the inhibition of IL-6-induced JAK2/STAT3 signaling in breast cancer cells through blocking of IL-6/IL-6R/gp130 complex formation.
In conclusion, we performed cell-based high-throughput screening using a Drosophila cell line and identified tubulosine as a strong inhibitor of JAK/STAT92E signaling in Drosophila cells. Furthermore, we showed that tubulosine inhibited IL-6-induced JAK2/STAT3 signaling by blocking IL-6Rα and gp130 binding. These results are the first demonstration of the mechanism of tubulosine action against cancer cells, and may suggest a potential therapeutic option for inflammation-associated diseases, including breast cancer.
 XML Download
XML Download