

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
The ovary is the main female organ that produces critical endocrine steroid hormones such as estrogen and progesterone. These endocrine hormones regulate the development of female sexual characteristics such as timing of reproductive cycles, breast development, and staging the uterus for successful pregnancy.
The ovary is also responsible for the development of mature oocytes capable of fertilization. The number of oocytes present in the ovaries at birth does not increase and determines the female reproductive lifespan [1]. At birth, approximately 1 million oocytes are present in women and approximately 10,000 are present in female mice [2]. The number of oocytes decreases during the development of female gonads and only approximately 400,000 oocytes remain at the puberty. Oocytes in the ovary become arrested in the dictyate stage of prophase in meiosis I before female is born [3]. When a woman's menstrual cycle starts and luteinizing hormone (LH) surge occurs, oocytes that have been arrested in prophase in the ovary enter meiotic resumption [4].
The oocyte is surrounded by pre-granulosa/granulosa cells, which together form the follicle as a functional unit in the ovary [56]. The smallest follicles are called primordial follicles (PFs) and their oocytes are surrounded with squamous pre-granulosa cells. They remain dormant inside the ovary until they receive activation signals which recruit them into the growing follicle pool.
Consequences of cancer therapy on ovarian follicles
Cancer therapy is like a 2-sided coin. It often causes many side effects in other organs including the risk of loss of ovarian follicles, while it increases the survival rate in the cancer patients [789]. As the ovarian follicles are the most sensitive cells, the exposure to the gonadotoxic agents such as chemotherapeutic agents, ionizing radiation, or environmental toxins consequently induce DNA damage and apoptosis in follicles. Thus, gonadotoxic agents eventually reduce the number of ovarian follicles, resulting in primary ovarian insufficiency in females [10]. Even though the follicles might survive the detrimental effects of gonadotoxic agents, the exposure can trigger genomic mutations in surviving follicles, potentially resulting in intergenerational effects [11].
Currently, there are options available for saving the ovarian follicles in cancer patients, which is dependent on their status [12]. Although ovarian tissue freezing is currently used as an option for future fertility for adult women and young girls before puberty, this procedure is not sufficient for long term maintenance of the endocrine system following the removal of the ovaries or depletion of the ovarian reserve [13]. Thus, proposed pharmacological approaches to prevent gonadotoxicity (Table 1) may be beneficial for prepubertal girls as they can protect ovarian follicles without surgery or delayed cancer therapy. However, a thorough understanding of the mechanism of ovarian follicle death by various cancer therapeutic agents is needed to prevent ovotoxicity and preserve fertility.
Table 1
The proposed mechanism of loss of primordial follicles (PFs) induced by chemotherapeutic agents and environmental toxins as well as the suggested fertoprotective adjuvants

| Proposed mechanism of loss of PFs | Proposed adjuvants | |
|---|---|---|
| Activation | Direct | AS101 [26], everolimus (RAD001) [27], INK128 [27], melatonin [29], ghrelin [29] |
| Indirect | AMH [31] | |
| Apoptosis | Direct | GnRH agonists [4041], GnRH antagonist [42], LH [46], ATM/ATR inhibitor [21], CHEK2 inhibitor [1924], and CK1 inhibitor [18], imatinib [48] |
| Indirect | S1P [5161], ceramide-1-Phosphate [52], zingerone [53], proanthocyanidin [54], gallic acid [55], mangafodipir [56], resveratrol [57], curcumin [58], capsaicin [58], erythropoietin [62], G-CSF [63], tamoxifen [64], dexrazoxane [656667] | |
AMH, anti-Müllerian hormone; GnRH, gonadotropin-releasing hormone; LH, luteinizing hormone; ATM, ataxia-telangiectasia mutated; ATR, ataxia telangiectasia and Rad3-related; CHEK2, checkpoint kinase 2; CK1, casein kinase 1; S1P, sphingosine-1-phosphate; G-CSF, granulocyte-colony stimulating factor.
![]()
Role of TAp63α in follicle death and molecular targets to limit TAp63α action
Oocytes from dormant primordial to early secondary follicle highly express tumor protein p63, TAp63α, an oocyte-specific isoform of p63 [1415]. TAp63α exists in the nucleus of oocyte as an inactive dimer in postnatal healthy oocytes and controls the fate of small follicles in the ovary [16]. However, DNA damage in oocytes induces the phosphorylation of TAp63α, leading to a conformational switch from an inactive dimer to an active tetrameric structure. This tetramer triggers apoptotic pathways through PUMA (BCL2 binding component 3; BBC3) and NOXA (phorbol-12-myristate-13-acetate-induced protein 1; PMAIP1) resulting in oocyte death [17]. Molecules upstream of TAp63α initiate the phosphorylation of TAp63α to make the tetrameric structure. Checkpoint kinase 2 (CHEK2) is an initiating kinase that phosphorylates dimeric TAp63α and recruits casein kinase 1 (CK1) to completely phosphorylate the primed TAp63α [181920]. The actions of CHEK2 and CK1 promote the formation of TAp63α tetramers, which are kinetically stable and irreversibly activate the apoptotic pathway. The chemotherapeutic agents, cisplatin and doxorubicin, have been shown to phosphorylate TAp63α. Thus, inhibitors of CHEK2 (BML-277 or CHEK2 inhibitor II hydrate) or CK1 (PF670462) block the tetramerization of TAp63α induced by cisplatin and doxorubicin and protect the ovarian reserve [1819]. Recent reports show that the use of CHEK2 inhibitors also protects the ovarian reserve from ionizing radiation [1921], suggesting that inhibition of TAp63α tetramerization through pharmacological intervention is a valuable approach to protect the ovarian reserve. These studies are based on understanding the mechanism of oocyte death by the treatment of gonadotoxic cancer therapies.
Loss of primordial follicles
Gonadotoxic therapies deplete PFs in the ovarian reserve. There are two prevailing theories regarding the effect of chemotherapeutic agents on the oocytes of dormant PFs (Table 1). The activation or burnout theory suggests that the chemotherapeutic agents activate dormant PFs into the growing follicle pool and proposes the use of inhibitors against the phosphatidylinositol-3-kinase (PI3K)/protein kinase B (AKT)/mammalian target of rapamycin (mTOR) pathway as an approach to preserve ovarian follicles from chemotherapeutic agents. In contrast, the inactivation or apoptosis theory suggests that chemotherapeutic agents directly destroy the oocytes of dormant PFs through activation of TAp63α and initiation of apoptosis. Thus, further investigation is required to identify and confirm the mechanisms by which chemotherapeutic agents induce death or activation of oocytes in dormant PFs. In order to effectively prevent the loss of oocytes, studies are needed to determine the specific cellular pathways that lead to oocyte/follicle loss based on the type and dose of chemotherapeutic agent as well as the duration of therapy.
Consequences of chemotherapeutic agents on oocytes of dormant primordial follicles: activation or apoptosis?
Intensive research has been conducted in the past decade to better understand how best to protect the ovarian germ cells from gonadotoxic agents such as chemotherapeutic agents or environmental toxins. However, it is unclear how ovarian germ cells die from chemotherapeutic agent insults and which approaches provide the best protection of the germ cells. Thus, this review revisits the current literature with a view towards unveiling the cytotoxic consequences of chemotherapeutic agents on PFs.
Loss of the ovarian reserve: activation
1. Direct activation
Existing literature supports the idea that activation of the PI3K/AKT/mTOR signaling pathway stimulates dormant PFs to enter the pool of growing follicles [222324]. Intriguingly chemotherapeutic agents such as cisplatin and cyclophosphamide are reported to induce PI3K/AKT/mTOR signals in oocytes without promoting the death of oocytes in PFs. Furthermore, Roness et al. [25] and Kalich-Philosoph et al. [26] demonstrated that cyclophosphamide induces PI3K/AKT/mTOR signals in the oocyte of the mouse ovary, causing the initiation of recruitment and growth of all dormant PFs (direct activation). This eventually results in the depletion of the ovarian reserve, and has been labeled as the follicle ‘burnout’ theory. Cyclophosphamide also induces death of growing follicles through apoptosis in granulosa cells, which results in a decrease of Anti-Müllerian hormone (AMH) and paradoxically results in the recruitment of dormant PFs into a cohort of growing follicles (indirect activation). Accordingly, AS101, an immunomodulator, decreased PI3K/AKT/mTOR signaling and eventually prevented the decline of follicles by follicle activation and loss from the treatment of cyclophosphamide. This study suggested that the dormant follicle activation through PI3K/AKT/mTOR signaling is a new mechanism of ovotoxicity underlying the loss of follicles induced by chemotherapeutic agents and environmental carcinogens. Thus, agents that suppress PI3K signaling, such as AS101, may be ideal adjuvants to preserve viability and numbers of dormant PFs when patients are treated with ovotoxins.
A study conducted by Goldman et al. [27] in 8-week-old C57BL/6J female mice supports the findings that cyclophosphamide promotes follicle activation through PI3K/AKT/mTOR signaling. In this report the ooplasm of dormant PFs highly expressed the downstream markers of mTOR signaling after the treatment with cyclophosphamide. Correspondingly, treatment with the mTORC1/2 inhibitors, everolimus (RAD001) or INK128, preserved the ovarian reserve and serum AMH levels. As a consequence, the mTOR inhibitors extended the fertility span in mice treated with cyclophosphamide.
A recent in vitro study by Lande et al. [28] that examined PFs in human ovarian tissues supports the activation theory. These investigators observed a statistically significant increase in the number of growing follicles in the human ovarian tissues following the treatment with high concentrations of 4-hydroperoxy cyclophosphamide (4-HC) or phosphamide mustard, which are cyclophosphamide metabolites, as compared with untreated ovarian tissues. Although there was no proposed mechanism in this report, the results correlate with others showing more developing follicles after the treatment with cyclophosphamide as compared to the untreated controls [26].
Jang et al. [29] supported the concept that cisplatin activates the PI3K/AKT/mTOR signaling pathway and stimulates dormant PFs. This study also provided evidence for melatonin and ghrelin being fertoprotective against cisplatin-induced ovarian damage by decreasing PI3K signaling. In the ovary, dormant PFs highly express Forkhead box O3a (FOXO3a) in the nuclei of oocytes [30]. A wave of follicle activation signals initiates PI3K/AKT signaling resulting in the phosphorylation of FOXO3a inside the oocyte nucleus. Phosphorylated FOXO3a shuttles out of the nucleus, stimulating activation of dormant PFs and converting the squamous pregranulosa cells into cuboidal granulosa cells (direct activation). In this report, the authors demonstrated that the treatment of mice with low doses of cisplatin (2.5 mg/kg) for 12 days activated dormant PFs through phosphorylation of FOXO3a. They also found that the treatment with melatonin and ghrelin synergistically suppressed the phosphorylation of FOXO3a by cisplatin and retained FOXO3a in the nucleus of oocytes of PFs. They further showed that the ovarian follicles express receptors for melatonin (MT1 and MT2) and ghrelin (GHSR1) in PFs [29]. These findings suggest that the dormant PFs may be protected from ovotoxins like cisplatin through high nuclear expression of FOXO3a. Furthermore, the combination of melatonin and ghrelin may serve as ovoprotectants/fertoprotectants to mitigate the adverse effects of cisplatin treatment.
2. Indirect activation
The indirect activation concept for loss of the ovarian reserve was recently highlighted by Kano et al. [31] demonstrating that AMH/Müllerian inhibiting substance (MIS) produced by granulosa cells protects the ovarian reserve from chemotherapeutic agents like carboplatin, doxorubicin or cyclophosphamide [32]. AMH/MIS plays an important role in the development of male and female gonads and the differentiation of the urogenital ridge. In females, AMH/MIS is mainly produced by granulosa cells from primary to medium sized antral follicles [33] and thus it is considered by the fertility clinics to be a biomarker for the ovarian reserve [34353637]. It serves an important role as a negative regulator of PF activation [3839]. The study by Kano et al. [31] demonstrated that co-treatment of AMH/MIS with chemotherapeutic agents protects PFs through suppression of PFs. According to this concept, chemotherapeutic agents destroy the growing follicles and thus reduce the production of AMH/MIS from them. Consequently, the lack of AMH/MIS production removes the suppressive effects allowing the PFs to be recruited into the pool of growing follicles (indirect activation). They also demonstrated that the treatment of AMH/MIS is reversible, showing that the cessation of treatment allows dormant PFs to resume ovarian folliculogenesis. Thus, co-treatment of AMH/MIS with chemotherapeutic agents is a therapeutic option for protecting the ovarian reserve.
Loss of the ovarian reserve: apoptosis
1. Direct apoptosis
Gonadotropin-releasing hormone (GnRH) agonists/antagonists are clinically used as ovoprotectants/fertoprotectants [404142]. Several reports show that the loss of ovarian cyclicity is prevented by co-treatment of GnRH agonist/antagonists with chemotherapeutic agents as compared with the chemotherapeutic agents alone [43]. Although the use of GnRH agonist has been promising, its usefulness has been debated due to the inconsistency of results in protection of ovarian follicles [1244]. A proposed mechanism indicates an increase in cAMP levels by GnRH agonist in oocytes leading to the suppression of oocyte apoptosis, although this finding was not confirmed in studies using mice and human [45]. This is thought to occur as a result of the passage of cAMP between oocytes and granulosa cells through gap junctions, eventually resulting in the survival of the ovarian follicles. The critical role of cAMP in ovoprotection is also supported by studies showing that treatment with LH provides protection against cisplatin. Research from Dr. Klinger's group showed that LH protects oocytes against cisplatin through the activation of the LH/choriogonadotropin receptor in ovarian cells [46]. In postnatal day 8 and adult mouse ovary, the LHCGR is expressed in cells surrounding follicles. Accordingly, exogenous 8-Bromo-cAMP and forskolin, factors mimicking the signaling pathway activated by LH, protect the ovarian follicles against cisplatin and initiate DNA repair kinetics in oocytes. Furthermore, LH stimulated the expression of KIT-ligand (Stem cell factor, SCF, KL, steel factor) and KIT (receptor), and protected oocytes against cisplatin-induced death in, both, oocytes and granulosa cells of the ovarian tissues in vitro. They proposed that protein kinase A activity, downstream of KL/KIT, protects oocytes from ovotoxicity by cisplatin followed by the activation of PI3K/AKT pathway to promote oocyte survival. However, it is not known whether the activation of PI3K/AKT pathway induced by LH treatment is necessary for protecting oocytes in PFs; in the case that it is needed, it must be on the level of threshold to promote oocyte survival.
Recent studies from our laboratory indicate that the chemotherapeutic agents directly destroy oocytes of dormant PFs and supports the inactivation concept. In support of the notion, cyclophosphamide treatment in three different mouse strains (C57BL/6J, CD-1 and BALB/cJ) did not promote any increase in the total number of primary and secondary follicles [47]. Our findings showed that the oocytes within the PFs are the most sensitive cells to DNA damage induced by cyclophosphamide treatment; this is in accordance with the previous data with cisplatin and X-rays [16]. Accordingly, oocytes of PFs were destroyed prior to damage to other ovarian cells without the activation of oocytes of dormant PFs, preserving the quiescent pregranulosa cells surrounding the oocytes of PFs. However, granulosa cells in growing follicles were also damaged. In addition, our results showed that the damaged granulosa cells are replaced within 3 days, if the follicles keep a portion of healthy granulosa cells, supported by proliferation of Ki67 markers and a subsequent increase in AMH production [47]. This suggests that the elevation of AMH does not represent the recruitment of dormant follicles into the pool of growing follicles. Furthermore, we found that the oocytes within the PFs presented temporal expression of phosphorylated-AKT as well as considerably prolonged expression of cleaved poly (ADP-ribose) polymerase (PARP) inside of the same PFs upon the treatment with cyclophosphamide or 4-HC, the active metabolite of cyclophosphamide in vivo. Moreover, cyclophosphamide induced time-dependent increases in apoptosis of oocytes of PFs as was evident by elevated expression of BAX and cleaved PARP. The oocytes expressed additional markers of apoptosis including γH2AX and phosphorylated TAp63α. Additional studies have demonstrated that cyclophosphamide-induced damage in oocytes of dormant PFs occurs via the p-ataxia telangiectasia and Rad3-related (ATR)>p-CHEK1/p-CHEK2>p-p63 apoptotic pathway [47], while cisplatin induces the p-ATR>p-CHEK2>p63/p73>PUMA/NOXA apoptotic pathway [172148]. The apoptotic pathway induced by cyclophosphamide eventually leads to the formation of TAp63α tetramers in the nucleus of oocytes, resulting in the induction of an irreversible apoptotic pathway. As shown in another report, ataxia-telangiectasia mutated (ATM), CHEK2, and CK1 inhibitors inhibit the tetramerization of TAp63α induced by cisplatin and doxorubicin [18], suggesting that the apoptotic inhibitors can block oocyte death at the molecular level [214849]. This suggests that PFs are destined to die rather than activate upon cyclophosphamide treatment, even though oocytes in PFs temporally present phosphorylated-AKT signals in response to chemotherapeutic agents. We assume that those follicles may contain genomic mutations or damaged DNA if the follicles with both the signals manage to survive. Taken together, these findings suggest that the apoptotic inhibitors instead of inhibitors of PI3K pathway should be used to protect dormant PFs from ovotoxins such as cisplatin, cyclophosphamide and X-ray [162147].
2. Indirect apoptosis by reactive oxygen species
Oxidative stress affects reproductive health in females through the production of reactive oxygen species (ROS). Although ROS play an important role in signal transduction during folliculogenesis, oocyte maturation, and embryogenesis [50], excessive ROS can induce damage to the ovary through the induction of granulosa cell death [51], follicle atresia or oocyte damage. In addition, it adversely affects the production of ovarian hormones and mature oocytes from healthy follicles. Of relevance to the preservation of fertility, reports suggest that the antioxidants such as S1P [51], ceramide-1-phosphate [52], zingerone [53], proanthocyanidin [54], gallic acid [55], mangafodipir [56], resveratrol [57], curcumin [58], and capsaicin [58] protect the ovary from ovotoxins including chemotherapeutic agents. Thus, oxidative stress-induced damage can be prevented by antioxidants through attenuating the free radical induced ovarian damages. However, there are only few reports that show direct evidences of ROS in oocytes of PFs. Thus, the effect of ROS on mitochondria in oocytes or granulosa cells should be further investigated [5960].
Conclusion
Understanding the exact mechanisms responsible for the disruption of the ovarian reserve induced by different classes of chemotherapeutic agents and environmental ovotoxins will allow for a rational approach to identify drug targets that protect oocytes and germ line integrity, select specific adjuvants and protect the ovarian reserve following cancer treatments (Fig. 1).
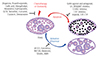 | Fig. 1Mechanism of primordial follicle (PF) loss by chemotherapeutics and ovotoxicity along with proposed fertoprotective agents to maintain the ovarian reserve. PFs consist of oocytes and squamous pregranulosa cells, while growing follicles are surrounded by cuboidal granulosa cells in the ovary. Fertoprotective agents have been proposed against the cytotoxic consequences of chemotherapeutic agents or ovotoxicity on PFs.
G-CSF, granulocyte-colony stimulating factor; GnRH, gonadotropin-releasing hormone; LH, luteinizing hormone; ATM, ataxia-telangiectasia mutated; ATR, ataxia telangiectasia and Rad3-related; CHEK2, checkpoint kinase 2; CK1, casein kinase 1; S1P, sphingosine-1-phosphate; C1P, ceramide-1-phosphate; AMH, anti-Müllerian hormone.
|
 XML Download
XML Download