

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Atopic dermatitis (AD) is a chronic inflammatory skin disease and the pathogenesis of AD is composed of elements of barrier dysfunction and alterations in cell-mediated immune responses12. Permeability barrier dysfunction is observed in lesional as well as non-lesional skin. The imbalance of T helper (Th) 2 to Th1 cytokines observed in AD can create alterations in cell-mediated immune responses. Skin fibrotic remodeling is also one of the major pathological features of AD, especially in its chronic phase3. Several studies have indicated that extracellular matrix components induce processes that result in the development of dermal fibrosis, and activate or protract the immune response. For example, periostin secreted from fibroblasts, an extracellular matrix protein induced by Th2 cytokines, promotes chronic allergic inflammation4. All of these appear to play roles in the development of AD. Although the pathophysiology of AD is characterized by structural abnormalities in the epidermis and immune dysregulation, the complex pathophysiology of AD is still not completely understood. In fact, incomplete efficacy of dupilumab5, clearly demonstrate complexity of the pathogenesis of AD in which a variety of factors other than Th2 cytokines should be involved.
The canonical Wnt/β-catenin signaling pathway is a fundamental mechanism accounting for various biological activities, such as cell proliferation, differentiation, and development. Levels of β-catenin are regulated by the Wnt/β-catenin pathway. In the inactive state, the β-catenin protein is degraded by a destruction complex composed of AXIN, glycogen synthase kinase 3β (GSK3β) and adenomatous polyposis coli (APC). On the activation of the Wnt/β catenin pathway, the AXIN GSK3β-APC complex is disrupted and GSK3β is inactivated, resulting in the protein stabilization and nuclear translocation of β-catenin. In the nucleus, β-catenin stimulates the transcription of target genes in cooperation with T-cell-factor/lymphoid enhancer-binding factor. To generate a transcriptionally active complex, β-catenin recruits the transcriptional coactivators, cyclic AMP response element-binding protein-binding protein (CBP) or its closely related homolog, p300, as well as other components of the basal transcription machinery, leading to the expression of a host of downstream target genes6. Aberrant Wnt/β-catenin signaling has been suggested to be involved in a variety of human pathologies, including cancers and metabolic, inflammatory, and fibrotic diseases7. ICG-001 selectively blocks the interaction of β-catenin with CBP. Recent studies have provided convincing evidence of the inhibitory effects of ICG-001 on disease models driven by Wnt signaling such as cancer8, organ fibrosis9, and acute lymphoblastic leukemia10, and, in particular, the improvement of asthma11. However, the effects of ICG-001 on AD have not been examined. Further, the mechanisms of ICG-001 in allergic disease are not yet fully understood. In this study, we investigated the efficacy of ICG-001 in a mouse model of AD-like dermatitis.
MATERIALS AND METHODS
Animals
Female hairless (Hr-/Kud) mice (8 weeks old, 20~23 g body weight) were purchased from Kyudo Company. (Fukuoka, Japan). The animals were group-housed, four mice per cage, under conventional conditions and had ad libitum access to a commercial diet and water. All experiments with mice were approved by the Ethics of Animal Experimentation Committee at Oita University (approval no. 1725002).
Development of oxazolone-induced AD-like dermatitis
Development of a hapten (oxazolone, Ox)-induced murine model with lots of features of AD (Ox-AD) was described in our previous studies1213. Animals were sensitized by two consecutive days of topical treatment with 50 µl of 5% Ox in acetone. After one week, mice were treated topically on both flanks with a total of 100 µl of 1% Ox in ethanol once every other day for an additional 3 weeks (total of 10 challenges).
ICG-001 administration and tissue preparation
ICG-001 was manufactured and provided by Prof. Kouji (Oita University, Japan). Mice were injected intraperitoneally with ICG-001 (4 mg/kg body weight) diluted in vehicle, i.e., dimethyl sulfoxide (DMSO), PEG300, Tween-80, and dH2O, or vehicle alone (vehicle control) once daily in an injection volume of 50 µl. On the last day of the experiment (day 29), the animals were sacrificed by cervical dislocation after transepidermal water loss (TEWL) and stratum corneum (SC) hydration were measured, and serum and skin samples were collected for the measurements and assessments described below.
Physiological assessments
TEWL and SC hydration on flanks were respectively measured using a Tewameter (TM300; Courage & Khazaka Electronic, Köln, Germany) and a Corneometer (CM825; Courage & Khazaka Electronic).
Histological analysis
Collected skin samples were fixed in 10% formalin and embedded in paraffin blocks. Tissue sections (5-µm thickness) were stained with hematoxylin-eosin for histological observation or Giemsa-stain for identification of mast cells, and then examined under light microscopy (Olympus, Tokyo, Japan). Epidermal and dermal thickness at six different locations on each image were measured as the distance from the basement membrane to the outer edge of the epidermis and from the epidermal-dermal junction to the dermal-subcutaneous fat junction, respectively, using a fluorescence microscope (BZ-X700; Keyence Corp., Osaka, Japan) at a magnification of 400× by an investigator blinded to the identity of the samples.
Immunohistochemistry
To detect myofibroblasts, skin sections were stained with a monoclonal antibody against α-smooth muscle actin (clone 1A4; Sigma-Aldrich, Taufkirchen, Germany). To examine whether CBP/β-catenin is involved in dermal fibrosis14, skin sections were stained with a monoclonal antibody against S100A4 (ab197896; Abcam, Cambridge, UK). Paraffin sections (5-µm thickness) were deparaffinized and incubated with a monoclonal antibody against α-smooth muscle actin and S100A4 overnight at 4℃. Detection was performed using an horseradish peroxidase (HRP)-conjugated secondary antibody (Dako envision+dual link system-HRP (diaminobenzidine+, DAB+); Dako, Carpinteria, CA, USA) or (Hystfine simple stain MAX-PO(R); Nichirei, Tokyo, Japan) followed by chromogenic detection using DAB (Dojin Chem., Kumamoto, Japan) as the substrate.
Counts of myofibroblasts and mast cells in the murine dermis
For quantification of myofibroblasts positive for α-smooth muscle actin, spindle-shaped cells in the dermis and mast cells in the Giemsa-stained dermis were counted in one or two randomly chosen high-power fields (magnification ×400) in each mouse by two experienced researchers in a blinded manner.
Cell culture
Culture of normal human keratinocytes (NHKs) (cat. no. 102-05; Cell Applications, Inc., San Diego, CA, USA) was performed as previously reported1516. When NHKs reached 70% to 90% confluence, expression of thymic stromal lymphopoietin (TSLP) was induced by stimulation with poly I:C (10 µg/ml), tumor necrosis factor (TNF)-α (20 ng/ml; R&D Systems, Minneapolis, MN, USA), and interleukin (IL)-4 (100 ng/ml; R&D Systems) with or without ICG-001 (5 µM). Hydrocortisone-free media were used during the stimulation protocol. The supernatants for enzyme-linked immunosorbent assay (ELISA) and NHKs for real-time polymerase chain reaction (PCR) were harvested 24 hours and 6 hours, respectively, after the stimulation.
Normal human dermal fibroblasts (NHDFs) (CSC-2F0; Cell Systems, Kirkland, WA, USA) were cultured in Dulbecco's modified eagle medium supplemented with 10% fetal bovine serum, 100 U/ml of penicillin, and 100 µg/ml of streptomycin at 37℃ in a humidified atmosphere of 5% CO2 and 95% air. The cells were cultured at an initial density of 5×104 cells/ml overnight. The medium was then replaced with serum-free medium and stimulated with human recombinant IL-4 (100 ng/ml; R&D Systems), IL-13 (100 ng/ml; R&D Systems) and with or without ICG-001 (5 µM) diluted in DMSO. The final concentration of DMSO was 0.1%. An equivalent concentration of DMSO (0.1%) was used as the vehicle medium in all experimental groups including the negative control. The supernatants for ELISA and NHDFs for real-time PCR were harvested 24 hours after stimulation.
mRNA isolation, reverse transcription, and real-time PCR
RNA isolation from skin samples minced in liquid nitrogen was performed using TRIzol reagent (Invitrogen, Karlsruhe, Germany) and the PureLink® RNA Mini Kit (Ambion, Thermo Fisher Scientific, Waltham, MA, USA). mRNA isolation from cultured NHKs and NHDFs was performed using the High Pure RNA Isolation Kit. mRNA was reverse transcribed to cDNA with Transcriptor First Strand cDNA Synthesis Kit (Roche Diagnostics, Basel, Switzerland). cDNA was analyzed by real-time PCR (The LightCycler® 96 System; Roche Diagnostics) using KAPA SYBR® FAST quantitative PCR master mix. Quantifications were normalized to the housekeeping gene, glyceraldehyde 3-phosphate dehydrogenase (GAPDH). Relative expression is given as the ratio between target gene expression and GAPDH expression. The primers used for real-time PCR are shown in Supplementary Table 1.
Measurements of levels of TSLP, thymus and activation-regulated chemokine, periostin and immunoglobulin E
Levels of TSLP, TARC, periostin and immunoglobulin E (IgE) in serum were examined by ELISA using a mouse TSLP Quantikine ELISA Kit (R&D Systems), a mouse CCL17/thymus and activation-regulated chemokine (TARC) Quantikine ELISA Kit (R&D Systems), a mouse periostin/OSF-2 Quantikine ELISA Kit (R&D Systems), and a mouse IgE quantitation kit (Yamasa Co., Ltd., Chiba, Japan) according to each manufacturer's instructions. Levels of TSLP and periostin in supernatant of cultured human keratinocytes or fibroblasts were determined by ELISA with human TSLP Quantikine ELISA Kit (R&D Systems) and human Periostin/OSF-2 DuoSet ELISA (R&D Systems) according to each manufacturer's instructions.
RESULTS
ICG-001 attenuated AD-like skin dermatitis and epidermal permeability barrier function in Ox-AD mice
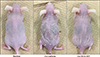
We examined the effect of ICG-001 in an AD-like model induced by repeated Ox treatments. ICG-001 or vehicle alone were injected intraperitoneally once daily (each group n=8). Forty-eight hours after the tenth Ox treatment to the sensitized mice, their skin appearance exhibited AD-like lesions such as erythema, edema, lichenification, and scale compared with the normal appearance of the non-treated control group (normal group n=4). ICG-001 attenuated Ox-AD skin lesions compared with vehicle group (Ox+vehicle; Fig. 1).
ICG-001 reduced epidermal hyperplasia, dermal fibrosis, and inflammatory cell infiltration in AD-like skin
Epidermal hyperplasia and skin fibrotic remodeling are major histopathological features in human AD. The application of ICG-001 suppressed epidermal hyperplasia and prevented the Ox-induced dermal fibrosis as assessed by the measurement of epidermal and dermal thickness and myofibroblast counts (Fig. 2A~D). Accordingly, S100A4 expression, which is upregulated by CBP/β-catenin14, was increased in Ox-induced dermal fibrosis, and this induction was reduced by the co-administration of ICG-001 (Supplementary Fig. 1).
AD-like skin showed marked dermal inflammatory cell infiltration. In particular, the numbers of mast cells in Giemsa stains were increased (Fig. 2E). ICG-001 treatment significantly suppressed the Ox-induced infiltration of mast cells.
Effects of ICG-001 on serum levels of TSLP, TARC, IgE, and periostin in Ox-AD mice
Levels of IgE and TSLP in serum were remarkably elevated in Ox-AD mice compared with control mice. Levels of TARC and periostin in serum also tended to be elevated in Ox-AD mice compared with control mice. ICG-001 treatment of AD mice suppressed serum levels of TSLP and TARC but not IgE. ICG-001 also tended to suppress serum levels of periostin, without statistical significance (Fig. 4). Moreover, mRNA expression of TSLP and periostin in lesional skin was not affected by ICG-001, as shown by real-time PCR in Supplementary Fig. 2.
ICG-001 does not affect the expression of TSLP in cultured human keratinocytes
TSLP expression induced by TNF-α, IL-4, and poly:IC in NHKs was not affected by ICG-001 (Supplementary Fig. 3A, B). These results indicate that ICG-001 does not have a direct effect on the expression and induction of TSLP in NHKs.
ICG-001 inhibits IL-4-, IL-13-induced periostin production by cultured human dermal fibroblasts
Next, we examined whether ICG-001 could inhibit IL-4, IL-13-induced periostin expression in NHDFs. Similar to our in vivo results, periostin mRNA expression was not affected by ICG-001, as shown by real-time PCR (Supplementary Fig. 3C). Furthermore, consistent with the observed trend in vivo, the level of periostin in the culture supernatant was significantly decreased, as shown by ELISA (Supplementary Fig. 3D), accompanied by a decrease in fibroblast counts (Supplementary Fig. 4).
DISCUSSION
The effects of ICG-001 on Ox-induced AD, namely decreases in TEWL, mast cell numbers in the dermis, epidermal and dermal thicknesses, serum levels of TARC and TSLP, and the increase in SC hydration, suggest that β-catenin/CBP-dependent signaling could be involved in permeability barrier dysfunction, allergic inflammation, and skin fibrotic remodeling in the pathogenesis of AD. Levels of S100A4 expression in skin were increased in Ox-induced AD and those were downregulated by the administration of ICG-001. These results suggest not only that β-catenin/CBP-dependent signaling is involved in the pathogenesis of Ox-induced AD but also that ICG-001 successfully inhibited the signaling in the present study. Further studies to check the involvement of β-catenin/CBP-dependent signaling in the pathogenesis of human AD should be also conducted in future.
ICG-001 is shown to have efficacy in various models driven by Wnt signaling, in particular organ fibrosis such as bleomycin-induced lung fibrosis or airway remodeling in asthma7. Therefore, the decrease in dermal thickness accompanying the decrease in dermal myofibroblast numbers might be a direct effect of ICG-001. Accordingly, ICG-001 inhibited increases in cultured human fibroblasts induced by IL-4 and IL-13 in the present study. However, mRNA expression of periostin in skin lesions and cultured human fibroblasts was not affected by ICG-001 treatment. Therefore, down-regulation of periostin in the supernatant of cultured fibroblasts and the tendency for decreased serum levels of periostin following ICG-001 treatment might be due to inhibition of fibroblast proliferation but not down-regulation of mRNA expression of periostin in fibroblasts. Furthermore, it was reported that upregulation of periostin mRNA expression by transforming growth factor (TGF)-β in cultured murine 3T3 fibroblasts was inhibited by ICG-00117, suggesting that fibroblasts treated with IL-4 and IL-13 might respond differently to ICG-1 compared to fibroblasts treated with TGF-β.
Accompanying the reduction of AD-like lesions, serum levels of TSLP were down-regulated by ICG-001 treatment. On the other hand, mRNA expression of TSLP in cultured human keratinocytes was not affected by ICG-001, suggesting that the decrease in serum levels of TSLP in ICG-001-treated mice was not a direct effect of ICG-001 on keratinocytes. Furthermore, mRNA expression of TSLP was not elevated in Ox-induced AD-like lesions, which were not affected by ICG-001 in the present study. It has been demonstrated that TSLP mRNA expression is induced transiently after stimulation18. Therefore, we speculate that skin samples harvested 48 hours after Ox treatment might not be suitable for evaluating TSLP mRNA expression.
In spite of the direct or secondary modulatory effects of ICG-001 on serum levels of periostin and TSLP, serum levels of IgE were not significantly affected by ICG-001. These findings contradict previous reports that periostin produced by fibroblasts acts on keratinocytes, inducing production of TSLP for Th2 skewing and, as a result, the cycle between Th2 and periostin maintains persistent inflammation in AD. A possible explanation underlying this discrepancy is presented below. Wnt signaling results in upregulation of SATB1 to recruit the β-catenin-p300 complex, which drives expression of GATA-3 in Th2 cells, with GATA-3 subsequently mediating the differentiation of Th2 cells19. As previously mentioned, ICG-001 inhibits the binding of β the binding of β-catenin to p300, so that ICG-001 might contribute to the promotion of Th2 responses. Meanwhile, the emergence of AD-like dermatitis was significantly prevented without alteration of serum IgE levels, indicating that IgE itself may not contribute to the pathogenesis of AD or suggesting that ICG-001 might target pathological pathways not related to Th2 in AD-like dermatitis.
The decrease in TEWL and the increase in SC hydration by ICG-001 treatment suggest that ICG-001 might directly contribute to normalizing permeability barrier function, although the roles of β-catenin/CBP-dependent signaling in permeability barrier homeostasis have not been elucidated. Moreover, possible effects of ICG-001 on a variety of cells including immunologic cells such as T lymphocytes remains unclear.
PRI-724, an isomer or enantiomer of ICG-001, has already been used in clinical trials for a variety of malignant hematopoietic diseases, cancers, and liver cirrhosis, demonstrating the safety and tolerance of PRI-724 without significant adverse effects2021. Taken together, β-catenin/CBP-dependent signaling might be contributed in the pathogenesis of AD, and could also serve as promising therapeutic targets especially by accounting for Th2 independent pathways. Further studies on human AD should be conducted for the therapeutic application of the blocking of β-catenin/CBP-dependent signaling.



 XML Download
XML Download