

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
The proportion of patients with estrogen receptor (ER)-positive breast cancer increased from 49.6% in 1996 to 70.0% in 2010 [1]. In patients with hormone receptor-positive (ER-positive and/or progesterone receptor; [PR] positive) breast cancer, antiestrogen therapy is the main treatment. The mechanism of action of tamoxifen is mediated by competitive binding to ER. Tamoxifen inhibits estrogen-related gene expression and blocks G1 phase of the cell cycle [2]. There was a randomized trial of tamoxifen for the treatment of early breast cancer. The proportion of recurrence for women with ER-positive breast cancer who used tamoxifen for 5 years was reduced by 50% (P < 0.01), and the mortality rate was reduced by 28%. On the other hand, in women with ER-negative breast cancer, the benefits of tamoxifen are unclear [3]. The antiestrogen therapy such as tamoxifen or an aromatase inhibitor lasting for 5 years reduces breast cancer recurrence and increases overall survival. Thus, tamoxifen has become the global standard of care for premenopausal patients with breast cancer. The recommendation of treatment takes into consideration that after 5 years, patients should receive additional antiestrogen therapy for 10 years [4]. However, adjuvant hormone therapy was discontinued for various reasons. Aiello Bowles et al. [5] reported discontinuation of hormone therapy and related adverse effects among patients with early breast cancer. The researchers reported that 18.2% of the patients had discontinued hormone therapy. Patients who discontinued treatment experienced adverse effects in many cases, with 41.8% reporting hormone/menopause-related symptoms.
Emodin (1,3,8-trihydroxy-6-methylanthraquinone) is a natural anthraquinone and phytoestrogen isolated from herbs [67]. Emodin has been reported to have antibacterial [8], anti-inflammatory [9], and anticancer effects [710]. Emodin can downregulate certain inflammatory markers, such as interleukin (IL)-1β, IL-6, IL-8, nuclear factor-kappa B, and tumor necrosis factor-alpha, and so on [11]. In addition, emodin has anti-inflammatory effects in several disorders, such as pancreatitis, arthritis, asthma, atherosclerosis, and glomerulonephritis [9]. The anticancer effect of emodin was reported for hepatocellular carcinoma cells [10], tongue squamous cancer cells [7], and pancreatic cancers [12] via apoptosis.
The aims of the present study were to investigate the combination effect of tamoxifen and emodin on ER-positive breast cancer cell lines and to explain the mechanism of the combination effect. In this study, we looked at patients with tamoxifen for passible material use to improve their hormone/menopause-related symptoms.
METHODS
Cell culture
MCF-7, T47D, and BT-474 breast cancer cell lines were purchased from the American Type Culture Collection (Manassas, VA, USA). The ZR-75-1 cell line was purchased from KOREAN CELL LINE BANK (KCLB; Seoul, Republic of Korea). Molecular classification of MCF-7 and T47D cell lines are “ER-positive and HER2 negative”. ZR-75-1 and BT-474 cell lines are “ER-positive and HER2 positive” [13]. The MCF-7 cell line was grown in Dulbecco's Modified Eagle's Medium (Welgene, Daegu, Republic of Korea), whereas ZR-75-1, T-47D, and BT-474 cell lines were grown in the Roswell Park Memorial Institute 1640 culture medium (Welgene, Daegu, Republic of Korea) with 10% of fetal bovine serum (Corning life sciences, USA).
Reagents
Antibodies against ERα and β-actin were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA) and those recognizing epidermal growth factor receptor (EGFR), cyclin D1, and phosphorylated extracellular signal-regulated kinase (p-ERK) were purchased from Cell Signaling Technology (Cell Signaling, Beverly, MA, USA). We used endoxifen instead of tamoxifen for our study to mimic the in vivo environment. Endoxifen is a therapeutically active metabolite of tamoxifen [14]. Endoxifen was purchased from Sigma Chemical Co. (St. Louis, MO, USA). Emodin, rhein (1,8-dihydroxy-3-carboxyl-9,10-anthraquinone) and chrysophanol (1,8-dihydroxy-3-methylanthraquinone) were purchased from Sigma-Aldrich.
Measurement of cell viability
Cell viability was measured using the EZ-Cytox cell viability assay kit (Itsbio, Seoul, Korea). The cells were incubated for 1 hour at 37℃ in a serum-free medium diluted with 1 kit reagent. Next, harvested cells resuspended in the media were carefully moved to empty 96- well plate and absorbance was measured using a microplate reader at 450 nm.
Isobologram analysis
Cell viability data were analyzed using in the CompuSyn software (ComboSyn, Inc., Paramus, NJ, USA). This software determines the combined effects of drugs: whether they are synergistic, antagonistic, or additive. The software was developed by Dr. Dorothy Chou in 2005. This software is based on the median-effect principle of the mass-action law [15]. In the isobologram graph, an antagonistic effect is above the oblique line (combination index [CI] > 1), a synergistic effect is below the oblique line (CI < 1), and an additive effect is on the line (CI = 1) [1516].
Western blot analysis
Cell lines were harvested and dissolved in radioimmunoprecipitation assay buffer (50 mM Tris-HCl pH 7.5, 150 mM NaCl, 1% Nonidet P40, 0.5% sodium deoxycholate, and 0.1% sodium dodecyl sulfate [SDS], supplemented with a protease inhibitor cocktail) (Gendepot, Katy, TX, USA). Equal amounts of protein (20–50 µg) were separated by SDS-polyacrylamide gel electrophoresis and transferred on to a nitrocellulose membrane. Membranes were blocked by incubating them with 2.5% skim milk for 1 hour and then incubated overnight with the appropriate primary antibodies (diluted 1:1,000), followed by a 1.5-hour reaction with the secondary antibody (diluted 1:10,000). Immunoreactive proteins were visualized by means of the enhanced-chemiluminescence reagent (Amersham Biosciences, Little Chalfont, UK).
Statistical analysis
All the results were compiled from a minimum of 3 independent experiments. Student t-test was performed to compare results of untreated (control) and treated group. Data were analyzed in the IBM SPSS ver. 18.0 (IBM Co., Armonk, NY, USA). Statistical significance was set to P < 0.05.
This study was exempted from review by the Institutional Review Board of the Konkuk University (study number: 7001355-201507-E-036).
RESULTS
Combined effects of endoxifen and emodin on human breast cancer cell lines
The ER-positive/HER2 negative; MCF-7, and ER-positive/HER2 positive; ZR 75-1, breast cancer cell lines were treated with a variety of concentrations of emodin or endoxifen for 48 hours. Cell viability findings are shown in Fig. 1A and B. The EZ-Cytox assay determined the cytotoxic concentrations of emodin and endoxifen (causing an approximately 20% reduction in cell viability) they were 60 and 4 µM, respectively. We decided to test combinations of concentrations of emodin (15, 30, 60 µM) and of endoxifen (2, 4 µM). Cell viability and microscopic findings for cells treated with the various combinations are shown in Fig. 2A and B. Emodin (60 µM) and endoxifen (4 µM) resulted in cell viability of 47.8% of MCF-7 cells after 48 hours (as compared to 60 µM Emodin alone: 56.5%, or 4 µM Endoxifen alone: 80.6%). In ZR-75-1 breast cancer cell lines, emodin (60 µM) and endoxifen (4 µM) yielded cell viability of 77.2% after 48 hours (as compared to 60 µM Emodin alone: 87.4%, 4 µM endoxifen alone: 86.6%).
The effects of the drug combinations were calculated in the CompuSyn software (Fig. 3A, B). In the MCF-7 breast cancer cell line, the CI value was 1.06586 between endoxifen (4 µM) and emodin (60 µM). In ZR-75-1 cells, the CI value was 1.20447 under the same conditions. Both CI values were >1, indicating an antagonistic effect between emodin and endoxifen in MCF-7 and ZR-75-1 breast cancer cells.
Subsequently, we validated the antagonistic effect in other breast cancer cell lines. The ER-positive/HER2 negative breast cancer cell line, T47D, showed similar antagonistic effects (Fig. 4A). Emodin (60 µM) and endoxifen (4 µM) yielded cell viability of 70.5% of T47D cells after 48 hours (as compared with 60 µM Emodin alone: 99.2%, or 4 µM endoxifen alone: 71.8%). The CI value was calculated and found to be 1.05637. The ER-positive/HER2 positive breast cancer cell line, BT-474, also manifested an antagonistic effect (Fig. 4B). Emodin (60 µM) and endoxifen (4 µM) yielded cell viability of 41% of BT474 cells after 48 hours (as compared with 60 µM emodin alone: 73.6%, or 4 µM endoxifen alone: 97.0%). The CI value was found to be 1.50427.
Analysis of the downstream mechanism of the antagonistic effect
To confirm the mechanism of the antagonistic effect, we performed western blotting assays. The MCF-7 and ZR-75-1 breast cancer cell lines were treated with emodin and endoxifen for 48 hours, and the harvested proteins were analyzed by western blotting. Treatment with the endoxifen and emodin combination; increased cyclin D1 and p-ERK amounts (Fig. 5).
Combination effects for similar chemical compounds on breast cancer cell lines
Subsequently, we studied the drug interactions between chemical compounds that are similar to emodin and endoxifen. Emodin, chrysophanol, and rhein are categorized as anthraquinones. The MCF-7 and ZR-75-1 breast cancer cell lines were treated with several concentrations of these compounds combined with endoxifen. Figs. 6 and 7 show cell viability and isobolograms for the drug interactions. The MCF-7 breast cancer cell line demonstrated antagonistic effects of the combinations of chrysophanol or rhein with endoxifen. The cell viability of MCF-7 cells was 86.2% when they were treated with endoxifen (4 µM) and chrysophanol (60 µM) for 48 hours (as compared with 60 µM chrysophanol alone: 88.0%, 4 µM endoxifen alone: 88.0%). The CI value was found to be 2.78039. The cell viability of MCF-7 cells was 95.4% when they were treated with endoxifen (4 µM) and rhein (60 µM) for 48 hours (as compared with 60 µM rhein alone: 87.8%, or 4 µM endoxifen alone: 89.9%). The CI value was calculated and found to be 3.09017.
Chrysophanol and rhein had antagonistic effects on ZR-75-1 breast cancer cell line. The cell viability of ZR75-1 cells was 86.7% when treated with endoxifen (4 µM) and chrysophanol (60 µM) for 48 hours (as compared with 60 µM chrysophanol alone: 97.7%, or 4 µM endoxifen alone: 87.5%). The CI value was found to be 1.63508. The combination of endoxifen (4 µM) and rhein (60 µM) yielded a cell viability of 97.2% of MCF-7 cells after 48 hours (as compared with 60 µM rhein alone: 96.5%, or 4 µM endoxifen alone: 88.6%). The CI value was calculated and found to be 3.72066.
DISCUSSION
The results of this study reveal antagonistic effects of emodin and endoxifen when used in combination against breast cancer cell lines. These antagonistic effects are associated with up-regulation of cyclin D1 and p-ERK. These results suggest that emodin, one of phytoestrogens, may interfere with the therapeutic effect of tamoxifen against ER-positive breast cancer cell lines.
In this study, we tested the drug combination on MCF-7 and ZR-75-1 breast cancer cell lines and validated the results in T47D and BT-474 breast cancer cell lines. The MCF-7 breast cancer cell line originated from the pleural effusion of a 69-year-old patient with metastatic breast cancer [17]. MCF-7 cells are widely used as an ER-positive cell line [13]. The T47D breast cancer cell line originated from the pleural effusion of a 54-year-old patient with metastatic breast cancer [18]. The T47D breast cancer cell line expresses high levels of PR. Thus, T47D cells are considered a major model for the research one progesterone in human cells [19]. The MCF-7 and T47D breast cancer cell lines are classified as luminal A (ER+, PR+/−, HER2−) [13]. The ZR-75-1 breast cancer cell line originated in the ascites of a 47-year-old patient with metastatic breast cancer [18] and is regarded as a luminal B (ER+/HER2+) subtype [13]. The BT-474 breast cancer cell line originated from a primary invasive ductal carcinoma of a 60-year-old patient with breast cancer [20] and is also a luminal B subtype (ER+/HER2+) [13]. In our study, MCF-7, T47D, ZR-75-1, and BT474 breast cancer cell lines all showed the antagonistic effects of our drug combination. According to these results, the combination of endoxifen and emodin has an antagonistic effect regardless of HER2 status.
Cyclin D1 forms a complex with cyclin-dependent kinase 4 (CDK4). The cyclin D1-CDK4 complex promotes S-phase entry by triggering E2F-dependent gene transcription [21]. In the western blotting analysis an increased cyclin D1 amount was observed in the cells treated with the drug combination. This result suggests that the drug combination promotes entry into the S-phase of the cell cycle. Cyclin D1 overexpression has been noted in breast carcinoma [21]. In the ZR-75-1 breast cancer cell line, EGFR expression increased in response to the drug combination. EGFR is composed of an extracellular region, a transmembrane segment, a juxtamembrane segment, a kinase domain, and sites of tyrosine phosphorylation. The extracellular region of EGFR undergoes dimerization, resulting in transmission of downstream signals [22]. The Ras–Raf–mitogen-activated protein kinase (MAPK) pathway, the phosphatidylinositol 3-kinase (PI3K)–Akt–mammalian target of rapamycin (mTOR) pathway, phospholipase Cγ pathway, the signal transducer and activator of transcription pathway, and the Src kinase pathway are all downstream effectors of EGFR [12]. EGFR plays an important role in many cancers, where it stimulates tumor growth, metastasis, and angiogenesis via these signaling cascades [23]. The Ras–Raf–MAPK pathway controls cell proliferation and survival. After phosphorylation of EGFR, EGFR forms a complex with proteins Grb2 and Sos, or interacts with them through the adaptor molecule Shc. This interaction recruits Ras-GDP and activates Ras-GTP. Ras-GTP activates Raf-1, and then stimulates Erk-1 and Erk-2. These 2 acts as transcription factors in the nucleus and ultimately promote cell proliferation and survival. The PI3K–Akt–mTOR pathway is associated with cell growth, apoptosis resistance, cell invasion, and cell migration [12]. PI3K phosphorylates phosphatidylinositol diphosphate to phosphatidylinositol triphosphate (PIP3). PIP3 activates the oncogene Akt. Akt inhibits tuberous sclerosis complex-2. As a result, Akt activates mTOR, which has effects on cell survival, cell proliferation, and metabolism [24]. In our study, increased p-ERK levels could explain the antagonistic effects of endoxifen and emodin on cell proliferation.
We performed drug interaction analyses between endoxifen and anthraquinone compounds similar to emodin (chrysophanol and rhein). Anthraquinones are benzoquinone derivatives that are synthesized by diverse pathways in plants and fungi. Anthraquinones have antimutagenic and anticarcinogenic activities. In addition, emodin, chrysophanol, and rhein have inhibitory effects on benzo[α]pyrene-mediated DNA damage in the HepG2 hepatoma cell line [25]. Nevertheless, in this study, emodin, chrysophanol, and rhein showed effects that are antagonistic to endoxifen's action on MCF-7 and ZR-75-1 breast cancer cell lines. Although we did not perform a mechanistic analysis of this finding, we hypothesized that the mechanism is similar to that of emodin.
Some investigators reported inhibitory effects of a combination of tamoxifen and genistein [2627]. Genistein is a phytoestrogen from soy that has been demonstrated to potentially inhibit the growth of breast cancer cell lines in vitro and to provide a relief of postmenopausal symptoms. Jones et al. showed that genistein inhibits tamoxifen's effects on the T47D breast cancer cell line. Tamoxifen (1.0 µM) decreased cell proliferation (92.8%) compared with the control (100%). By contrast, the combined treatment with genistein and tamoxifen improved proliferation (96.7%) of the T47D breast cancer cell line. This result was interpreted as a reversal of cell cycle arrest: Tamoxifen induces G1 arrest; however, the combined treatment with genistein and tamoxifen reverses the G1 arrest, thereby normalizing the cell cycle [26]. Ju et al. [27] determined whether genistein negates the inhibitory effect of tamoxifen on the growth of MCF-7 breast cancer cells in nude mice. They studied the effect of the drug under several conditions and showed antagonistic effects. The combination treatment with genistein and tamoxifen resulted in a significantly larger tumor surface area compared with tamoxifen alone (2.5 TEG, 75.1 mm2; 5TEG, 50.9 mm2; 2.5TE, 14.4 mm2; 5TE, 13.7 mm2). This study revealed that the combination treatment recovered pS2 expression via cyclin D1 overexpression as compared to the treatment with tamoxifen alone. Thus, these 2 studies verified the antagonistic effect between tamoxifen and other phytoestrogens, in agreement with our results.
In conclusion, our data indicate that the combination treatment with endoxifen and emodin has an antagonistic effect via cyclin D1 and pERK overexpression in ER-positive breast cancer cell lines. These results suggest that emodin may attenuate tamoxifen's therapeutic effects. Therefore, patients with ER-positive breast cancer should be careful when consuming phytoestrogens.
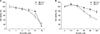






 XML Download
XML Download