

 PDF
PDF Citation
Citation Print
Print



Abbreviations
AHR
airway hyper-responsiveness
AM
alveolar macrophages
APC
antigen-presenting cell
AR
airway reactivity
BAL
bronchoalveolar lavage
COPD
chronic obstructive pulmonary disease
DAMP
damage-associated molecular pattern
DC
dendritic cell
GC
glucocorticoids
HDM
house dust mite
ICAM-1
intercellular adhesion molecule-1
ICS
inhaled corticosteroids
ILC
innate lymphoid cell
IM
interstitial macrophages
iNOS
inducible nitric oxide synthase
IRF
IFN regulatory factor
LABA
long-acting β2 agonist
LPG
laboratory of genetics and physiology
MDA
melanoma differentiation-associated gene
NLR
Nod-like receptor
NSAIDs
non-steroidal anti-inflammatory drugs
OVA
chicken ovalbumin
PAMP
pathogen-associated molecular pattern
PRR
pattern recognition receptor
RIG
retinoic acid-inducible gene
RLR
retinoic acid-inducible gene-like receptor
RSV
respiratory syncytial virus
RV
rhinovirus
INTRODUCTION
Asthma is a frequent health concern globally associated with significant morbidity, hospitalization costs, and even fatal prognosis. Asthma is defined as a heterogeneous disease usually characterized by chronic airway inflammation (1) affecting almost 334 million people worldwide including up to 10% of adults and 30% of children, with an expected increase to around 400 million by the year 2025 (2). The heterogeneity of asthma depends on numerous risk factors, determinants, degree of airflow obstruction, hyper-responsiveness, severity and type of airway inflammation (3). Environmental triggers including allergens and pollen in the air and the diet, air pollutants, tobacco smoke, genetic differences, and respiratory viruses may exacerbate asthma, and play a vital role in worsening the outcomes (4). Approximately 100 major or minor genes may be associated with the development of asthma mediated via innate immunity, Th2 effector function, mucosal and lung functions along with airway remodeling (5). The immune responses and possible treatment strategies may be clear in classical (Th2 signature) asthma. However, in cases of severe and refractory asthma the multidimensional mechanisms and related immune response have yet to be defined. In non-Th2 response, the severity of augmented asthma and increased airway remodeling associated with poor prognosis have been correlated with a complex innate and adaptive immune response (6). Herein, the innate and adaptive immune signatures of asthma exacerbated by respiratory viral infection are discussed.
ASTHMA EXACERBATION
Asthma exacerbation is characterized by higher and intense asthma severity, increased airway remodeling, a lower response to bronchodilator and anti-inflammatory treatments, resulting in pulmonary dysfunction. As a phenotypically heterogeneous disease, asthma involves multifaceted interactions between multiple risk factors and disease determinants (14). The diversity of asthma may be attributed to a complex interplay between risk factors (either intrinsic or extrinsic) and the specific disease determinants (viruses, bacteria, etc.). Rhinovirus (RV) is the major and most frequent determinant than respiratory syncytial virus (RSV), influenza, other corona viruses, or even pneumonia-inducing bacteria (7). Hospitalized patients manifesting asthma features carry a history of respiratory viral infections (mainly RVs) along with major extrinsic factors such as cigarette smoking, age, obesity, rhinosinusitis, pneumonic symptoms, and intolerance to non-steroidal anti-inflammatory drugs (NSAIDs) (8). The intrinsic risk factors (genetic polymorphisms, enhanced airway epithelial lysis, and apoptosis) are strongly correlated with exacerbation in asthmatic hosts, and exaggerated virus replication in infected cells may result in reduced innate immune response, and concomitant reduction in the expression of innate interferons (types I, II and III) (910). This subsection will highlight the etiology, the pathophysiological mechanism and ongoing treatment strategies targeting exacerbated asthma.
Respiratory virus infections inducing asthma exacerbation
In addition to classical allergic asthma and delayed immune response, respiratory viral infections involved nearly 80% of cases diagnosed with hospitalized asthma exacerbation (11), including pediatric cases constituting nearly 63% of them (12). Among the respiratory viruses, RVs represent the most prevalent viral pathogens responsible for around 60% of virus-associated asthma exacerbations (713). Different murine models of experimental RV infection including chronic obstructive pulmonary disease (COPD), airway hyper-responsiveness (AHR) to histamine and subsequent lung inflammation were used to demonstrate the typical features of asthma (714). Emerging evidence supporting asthma exacerbation suggests a deficiency in antiviral immunity and loss of epithelial barrier integrity in individuals prone to severe refractory asthma (15). Although the exact mechanisms underlying asthma exacerbations have yet to be defined, studies suggest that RV infection of airway epithelium leads to cytokine interaction with allergic inflammation to overwhelm both innate and antigen-specific Th2 pathways resulting in enhanced Th2-related inflammation (71617). Moreover, both in children and adults, a complex association between RV infection and asthma exacerbation may also induce a non-Th2 immune response, along with increased AHR, mucus hypersecretion, airway remodeling, and respiratory failure (6).
Clinical pathophysiology of asthma exacerbation
RV-induced asthma is not only confined to the upper airway, but conclusive clinical evidence suggests that it is very common in the lower respiratory tract of children along with critical pneumonia signatures (814). In the respiratory airways, mucosal lining and cilia eventually trap airborne antigens and other harmful particles to prevent entry into the lung via muco-ciliary transport. Airway epithelium is exposed to foreign antigens, resulting in a cascade of secretory mechanisms such as mucus secretion, followed by secondary defense mechanisms mediated via cytokines and chemokines (18). The most cardinal features of asthma include inflammatory cellular infiltration, and mucus hyper-secretion along with AHR. Upon alveolar injury or pathogen invasion, the levels of patrolling immune cells, antigen-presenting cells (APCs), alveolar macrophages, and dendritic cells (DCs) are persistently elevated in conjunction with airway epithelium where leukocyte infiltration resolves subsequent inflammation (Fig. 1). The immune response starts with the activation of T lymphocytes by APCs, along with concomitant interaction between infiltrated cells, cytokines, chemokines and related mediators triggering and maintaining asthmatic airways (19). Differentiation of Th0 cells into Th1, Th2, Th17, Th22, and Th9 cells is a complex phenomenon and poorly understood. Classical or eosinophilic asthma may exhibit a ‘high Th2’ molecular phenotype, and elevated serum IgE level indicating type 2 allergic mechanisms (20). The Th2 airway inflammation is characterized by eosinophilia and exhaled nitric oxide exacerbating the risk of asthma (21). By contrast, asthma exacerbation was characterized by Th1 response with neutrophil infiltration or even concurrent Th1-Th2-Th17 expression (22). Non-Th2 asthmatic pathophysiology has been linked to dysregulated innate immunity, including the activation of inflammasome pathways and development of airway neutrophilia (23). Recent studies investigating the mechanism of asthma exacerbation revealed a positive association between respiratory viral infections and activation of microRNAs and inflammasome-activating genes, and enhanced corticosteroid resistance (2425)
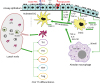 | Figure 1Interplay between innate immune cells in airways for adaptive immunity. The innate immune response in airway epithelium is initiated by contact with innocuous antigens, which triggers the expression of PRRs in lung epithelium. The activation of innate signaling receptors in airway epithelium results in migration of imDC to the mucosal epithelium via chemokine secretion from the epithelium. Following antigen stimulation, the AM are also activated to secrete GM-CSF, IL-8, and TNF-α that facilitate the activation of DCs. Moreover, epithelial cells release IL-1β and TNF-α, which induce the release of innate cytokines GM-CSF, IL-17E, TSLP, and IL-33 via binding to their respective receptors on the epithelium. The activation of airway DCs by the combined streaming of cytokines secreted by the epithelium promotes migration into draining lymph nodes, and induction of adaptive Th response.imDC, immature dendritic cells.
|
Treatments for asthma exacerbation
The efficacy of specific treatments and therapeutic agents targeting asthma has yet to be clearly defined. Most of the therapies are symptom-oriented and palliative based on severity. In general, current approaches include treatment with inhaled corticosteroids (ICS), long-acting β2 agonists, leukotriene receptor antagonists, inhaled IFNs, and anti-IgE therapies. Glucocorticoids (GC) are practicing as a mainstay to treat airway inflammatory disorders like rhinitis, chronic sinusitis, COPD and asthma. In addition to displaying well-known anti-inflammatory functions, GC is found to spare or enhance local innate response in respiratory epithelium (26). Treatment with anti-IL-5 (mepolizumab), anti-IL-13 (lebrikizumab), and even anti-IL4-Rα (dupilumab) has been shown to reduce the levels of eosinophils and asthma exacerbation characterized by Th2 features (27). Generally, treatment of allergic airway inflammation, AHR, and respiratory virus-induced asthma with corticosteroids is ineffective, especially in RV-induced asthma exacerbation associated with neutrophil infiltration (11). Nearly 50% of cases showing exacerbated asthma involving neutrophil infiltration are not resolved with corticosteroid therapies (28). The etiology of corticosteroid-resistant asthma was strongly related to airway epithelial resistance via NF-κB and JAK activation, oxidative stress, and limited activity of transcriptional corepressor histone deacetylase-2 (29). Moreover, the ssDNA-like antisense agents including peptide-conjugated phosphorodiamidate oligomers (pleconaril and morpholino) are used as effective anti-enterovirus therapies against RVs by targeting highly conserved internal ribosome entry sites (30). Besides, inhaled IFNs including type I IFN (9), particularly IFN-β (31) were used to alleviate respiratory virus-induced exacerbation of asthma symptoms (32). Therefore, appropriate therapies are urgently needed to manage asthma associated with non-eosinophilic or neutrophil-eosinophil mixed allergic asthma responses. Furthermore, corticosteroids diminish the response to CD4+ Th9 cells in allergic asthma. The Th2-mediated asthma was controlled using corticosteroids via inhibition of the epithelial TARK (CCR4 ligand expressed by Th2 cells) (33). However, Th9 cells express functional CCR3, CCR6, and CXCR3, and corticosteroids may enhance CXCR3 ligand (IP-10) in asthmatic lungs, resulting in inadequate control of Th9-mediated asthma (34).
SMALL ANIMAL MODELS FOR ASTHMA EXACERBATION INDUCED BY RESPIRATORY VIRAL INFECTION
Due to the logical limitations of human disease models, numerous small animal models, in vitro models, and mathematical equations have been widely used to study asthma pathogenesis. These models may have widened the scope of investigation into the mechanistic pathways of asthma pathogenesis, and subsequent therapeutic discovery. Therefore, even though these models have some limitations, the hypothesis can be proved by analyzing all the models collectively and integrating their outcomes. In particular, respiratory virus-induced asthma exacerbation is a big concern, due to its clinical sequelae, and the intricate relationship between pathogenicity of respiratory viruses and defense mechanisms of model animals. Development of asthma models depends on the small animals (mice, rabbits, etc.) used and the viruses (RV, RSV, influenza, etc.) investigated. It also depends on the strain variation along with pre-exposure allergen sensitization (chicken ovalbumin [OVA], house dust mite [HDM], etc.). The results are similar to acute or chronic asthma models. Influenza virus infections in HDM- or OVA-sensitized mice usually induce critical features of asthma, whereas influenza and RSV models may fail occasionally due to the overwhelming cytokine response and canonical eosinophilic trafficking (35). Further, murine models, particularly those involving allergic inflammation (OVA, HDM) or RV-induced exacerbations are associated with varying levels of advantages and disadvantages from a scientific perspective (36). Many murine models of asthma associated with RV infection have failed, probably due to RV serotypes, a majority (90%) of which binds to human intercellular adhesion molecule-1 (ICAM-1) but not the mouse ICAM-1. By contrast, 10% of RVs such as RV-1B use different adhesion molecules and bind to the mouse low density lipoprotein receptors (17). Most of the murine models show inflammatory changes and typical asthma features resembling human bronchial obstruction, AHR, epithelial hypertrophy, goblet cell metaplasia, infiltration of innate cells, trafficking of Th cells and concurrent cytokine-chemokine interactions (3738). Basically, models of acute and chronic inflammation are highly specific to allergens (OVA, HDM, cockroach extract, etc.), murine strains, sensitization, and virus exacerbation protocols used (39).
INTERPLAY BETWEEN HOST IMMUNITY AND RESPIRATORY VIRAL INFECTION IN ASTHMA EXACERBATION
Respiratory viral infections linked to asthma pathogenesis may differ in outcomes, depending on the related risk factors. The common cold virus, RV, has emerged as the primary etiological agent associated with exacerbations and other pathological outcomes of asthma. After the onset of RV attacks, the typical or exacerbated asthma features develop following the activation of different innate responses. The critical pathogenesis of RV-exacerbated asthma is triggered by impaired respiratory epithelial function inducing the release of variable proinflammatory mediators to attract airway inflammatory cells like eosinophils, neutrophils, macrophages, and lymphocytes (40).
Role of innate immunity in asthma exacerbation caused by respiratory viral infection
The host innate immune response is characterized by critical mechanisms for prompt detection, binding, and elimination of invading pathogens. The innate immune system is fortified with various pattern recognition receptors (PRRs) such as TLRs, retinoic acid-inducible gene (RIG)-like receptors (RLRs), and Nod-like receptors (NLRs) for recognition and response to pathogen-associated molecular patterns (PAMPs). Here we will focus on the mechanisms of different PRRs against respiratory viruses, and innate cells to orchestrate innate immune response in asthma development.
PRRs in allergic airway inflammation
Recognition of allergens or respiratory viral infections (such as RVs) by PRRs is followed by enhanced secretion of inflammatory cytokines including types I and III IFNs (41). RV infection-increased numbers of subepithelial IFN/PRR-expressing inflammatory cells are related to RV loads in airways and concurrent severity of asthma (42). TLRs are transmembrane and noncatalytic PRRs expressed on macrophages, DCs, fibroblasts, epithelial or other non-immune cells (43). The initial innate responses are regulated by TLRs via activation of TRIF and MyD88 signaling pathways, ensuring the synthesis of inflammatory mediators via NF-κB pathway (44). The asthmatic conditions associated with neutrophilic trafficking are characterized by the increased threshold of IL-8 and IL-1β, and upregulation of TLR2 and TLR4 via MyD88 and TRIF pathways followed by NF-κB and MAPK signal activation. Besides, TLR2 and TLR4 are associated with lung dysfunction, and pediatric asthma (45). Following the initiation of TLR4 activation, pro-inflammatory cytokines are released to attract innate cells (macrophages, mast cells and neutrophils), and eventually leading to ROS production (46). TLR7 activation is strongly associated with the induction of Th1 axis, masking the development of classical Th2 immunity, and IgE assembly in response to allergen-induced airway inflammation or RV-exacerbated asthma (47). The RLR family comprises RIG-I, melanoma differentiation-associated gene (MDA) 5, and laboratory of genetics and physiology (LPG), which have been shown to mediate the innate immune response against double-stranded RNA (48). The RLR-induced signaling results in a robust production of type I IFN via activation of NF-κB, MAP kinase and IFN regulatory factor (48). Furthermore, RLRs (RIG-I and MDA5), which could be induced initially through TLR3/TRIF pathway, show the coordinated role with TLR3 in exhibiting innate defense to RV (41).
Currently, asthmatic features are highly linked to NLR-dependent maturation and secretion of pro-IL-1β and IL-18. NLRs can be activated by physical damage to the plasma membrane induced by respiratory viruses and host-derived damage-associated molecular patterns (DAMPs), which stimulates the caspase 1–activating platform (inflammasome) that plays a vital role in host defense against RV (2549). RV infection activates NLRP3 and NLRC5, which leads to the activation of caspase 1 and the hyper-production of IL-1β (50). Thus, in response to allergens or respiratory virus infections, the PRRs (TLRs, RLRs, and NLRs) play a crucial role in the recognition of viruses, and subsequent induction of innate response mediated via inflammatory cytokines and stimulation of IFNs.
Innate response of airway epithelial cells
Airway epithelium, the first-line barrier to external environment and maintenance of internal milieu, expresses multiple PRRs for rapid detection and response via release of microbial PAMPs or DAMPs under cellular stress, tissue damage, necrosis, and cellular death (551). Airway epithelium detects hRV infection via TLRs, RIG-I and MDA5, followed by activation of signaling pathways that lead to the synthesis of pro-inflammatory cytokines and type I IFNs (4152). Upon RV infection, airway epithelium may express type I IFN-β and type III IFN-λ resulting in defective epithelial apoptosis during exacerbation (91053). Airway epithelium orchestrates and triggers airway inflammation via release of CCL2 and CCL20 in response to allergen stimuli. The airway epithelium activates the immunomodulatory role of DCs via release of multiple cytokines (TSLP, IL-25, IL-33, SCF, GM-CSF, etc.) following activation of TLRs in response to external stimuli and allergens (54). Simultaneous activation of DCs may induce the release of CCL17 and CCL22 mediating Th2 response via CCR4, followed by the synthesis of RANTES, Eotaxin-1, and Eotaxin-2 to attract eosinophils via CCR3 activation pathway. Specifically, TSLP promotes growth and differentiation of basophils to induce IgE class switching controlled by IL-4; however, IL-17E (IL-25) and IL-33 may activate nuocytes to induce simultaneous expression of IL-5 and IL-13 for eosinophilic trafficking, goblet cell metaplasia, and AHR development triggered by the virus (55). In addition to cytokine synthesis, the respiratory epithelium may produce DAMPs (ATP, uric acid, ROS, etc.) to regulate the immune response in atopic conditions or even a single exposure to allergens due to activation of DCs and other innate cells via NF-κB signaling pathway (56). Moreover, in response to antigen exposure, the airway epithelium secretes prostaglandin E2 to diminish DC reactivity by binding to the EP4 receptor, and thus facilitate the suppression of airway inflammation (57).
Eosinophils
In allergic inflammation and respiratory virus-induced asthma, the infiltration of key innate cells including eosinophils and neutrophils explains asthma pathogenesis individually or via mixed immune response (58). Eosinophils play an innate antiviral role via production and secretion of neurotoxin, ROS, and eosinophilic cationic proteins, cytokines and lipid mediators. Thus, eosinophils exhibit multiple roles such as epithelial barrier function, intrinsic interaction between innate and adaptive immunity, and tissue remodeling (59). Following RV infection, the infiltrated eosinophils also secrete cationic proteins and leukotrienes, thereby resulting in AHR upon activation of eosinophils following the secretion of RANTES by the airway epithelium (15). However, increased levels of nasal or BAL eosinophils are related to allergic airway inflammation, and respiratory viral infections triggering either acute or chronic asthma (17383960). In human respiratory infections, and various murine models of asthma, the eosinophils act as APCs to activate naïve CD4+ Th cells via surface markers MHC-II and CD40 in response to stimulation by cytokines including IFN-γ, GM-CSF, IL-3, IL-4, and IL-5 (61).
Neutrophils
Airway neutrophilic trafficking occurs early as a first-line defense after the onset of respiratory virus-infections and concomitant asthma exacerbation (52). Neutrophils release chemotactic factors and granular proteins to activate monocytes and macrophages in the airway and induce an innate response (62). Despite the innate host defense, the activated neutrophils exhibit immunoregulatory roles to secrete ROS, dsDNA and elastase containing neutrophil extracellular traps (NETs) via programed cellular death known as NETosis (63). In adults, the exacerbation of virus-induced asthma is characterized by an influx of sputum neutrophils and neutrophil elastase, responsible for airway obstruction, mucus secretion, and lower respiratory tract symptoms (64). Moreover, the neutrophil-mediated dsDNA may enhance the expression of Th2 cytokines (IL-4, IL-5, and IL-13) in nasal or BAL fluid, which mediate the classical type 2 immune response (6063). In acute asthma exacerbations by RVs, neutrophil infiltration occurs most predominantly in sputum or lungs, and is strongly induced by IL-8 and G-CSF (17). Several murine models suggest classical and neutrophil-eosinophil interaction in BAL and inflamed asthmatic lungs (71438); however, the exact mechanism underlying the role of neutrophils in asthma has yet to be established.
Macrophages
Macrophages, DCs, and monocytes constitute the mononuclear phagocytic network in the host innate response to airborne antigens. The macrophages are the most abundant innate immune cells in the airway lumen, and are involved in the early phase of host defense against RV exposure (37). In asthma pathogenesis, macrophages play a vital role in coordinating adaptive immunity, regulation of airway inflammation, and subsequent tissue repair (52). In early allergy response, the influx of very short-lived proinflammatory monocytes, interstitial macrophages (IM) and airway macrophages (AM) appears to mediate early- and late-phase activation and differentiation (65). The innate response of resident AM is initiated by the secretion of IL-10, and IL-12 (anti-Th2) cytokines (66). In contrast, the IM regulates the DC function via IL-10 followed by neutrophil infiltration and subsequent attenuation of Th2 response to allergen exposure and other determinants of airway inflammation (66). RV infection also induces the release of IFN-α from AM to maintain the antiviral status of the airway epithelium and facilitate homeostasis of the lung (67). Thus, the allergen-primed macrophages play a protective role against tissue damage, and the asthmatic mice show increased allergic inflammation and AHR upon depletion of those macrophages. Several in vivo murine and ex vivo models corroborated the intense induction of CCL11 and IL-4 expression in the AM via phenotypic changes in response to allergen and RV exposures (37). The classical (M1) polarized macrophages utilize transcription factors STAT1/NF-kB, and are activated via IFN-γ, GM-CSF, LPS, to secrete IL-1, IL-8, IL-12p40, inducible nitric oxide synthase (iNOS), IFN-γ, TNF-α, and MIP-1α. In contrast, the M2-polarized macrophages express a different set of phagocytic receptors, and are activated by M-CSF, IL-4, IL-8, and IL-13 resulting in the secretion of arginase, IL-10, and TGF-β, using the transcription factor STAT3/STAT6 (68). A few murine and human models also demonstrated the sequential differentiation of M2-polarized macrophages in both RV-induced acute and chronic asthma via CCL11, and CCL17-dependent eosinophilic expression (3769).
γδ-T cells
In asthma, the γδT cells are strongly associated with epithelial tissue homeostasis, modulating both antiviral innate and cell-mediated immune responses. They facilitate a neutrophil-mediated host defense mechanism against respiratory infection (70). The murine asthma models were used to demonstrate the role of γδT cells in AHR, and prime the antiviral innate response to RV-induced asthma exacerbation (71). Moreover, in animal models of OVA-induced allergic asthma, the γδTreg associated with immunosuppressive function express both Foxp3 and TCRγδ (72). In humans, upon RV or allergen exposure, the critical features of asthma such as increased viral load, airway obstruction, AHR, and exaggerated eosinophilia in BAL, are strongly associated with γδT-cell population in the airway and the blood (71). In the murine asthma model, the γδT cells attenuate allergic airway inflammation. By contrast, a few reports suggested the insignificant effect of anti-γδTCR treatment on pulmonary viral load, IFN-β response, and RV-specific serum IgG (71). Therefore, multidimensional ex vivo and in vivo models are needed to elucidate the immunoregulatory roles of γδT-cells.
Innate lymphoid cells (ILCs)
In airway mucosal tissues, ILCs play a critical role in host defense and tissue homeostasis, and facilitate the interaction between innate cellular signals and adaptive immune response (73). Based on the response to innate cellular signals (TSLP, IL-17A, IL-33, IL-1β, and TGF-β) generated by the asthmatic airway epithelium, ILCs can be subdivided into ILCs-1, 2, and 3, depending on the cytokines expressed (74). Cytokine synthesis in ILCs may be linked to macrophage polarization (M1 or M2) to orchestrate asthma pathogenesis (75). The roles of ILCs in asthmatic airways vary depending on the type of differentiation, cytokine production, and asthma pathogenesis. Usually, ILCs express and secrete high levels of Th1, Th2 and Th17 cytokines, whereas ILC2 shows an eosinophilic Th2 asthmatic signature via induction of IL-4, IL-5, IL-9, and IL-13 in response to IL-25 and IL-33 secreted by the asthmatic epithelium (76). However, ILC1 and ILC3 coordinate with other innate cells, or neutrophils although a few reports suggested that ILC1 secretes IFN-γ via T-bet as an innate complement to CD4+ Th1 cells (77). Besides, in some animal and human models, ILC3 secretes IL-17A, TNF-α, and IL-22 to exacerbate asthma pathogenesis (78).
Role of IFNs in innate immunity during asthma
The innate IFNs in asthma boost host antiviral defenses, resulting in reduced RV load and RV-induced acute exacerbation, although the precise mechanisms are unknown. Based on the tropism for different receptors, the type I IFNs consist of a single IFN-β, and 14 IFN-α subtypes; the type II includes the IFN-γ subtype, and the type III IFNs comprise IFN-λ 1, 2, 3, and 4 subtypes. Type I and III IFNs share a similar role in innate anti-viral immunity (79). Usually, airway epithelium and hematopoietic cells produce type I and III IFNs, whereas type II IFN (IFN-γ) secretion is restricted to Th1, NKT, γδT cells, macrophages, DCs, and NK cells.
Type I IFNs
Notably, type I IFNs are only induced after RV exposure to airway epithelium or in vitro stimulation of coherent cells with either type I IFNs or specific TLR agonists. The type I IFNs signal via IFNAR in conjunction with ISG induction to yield an anti-RV phenotype (80). Type I (α/β) IFNs in airway epithelium or sub-epithelial innate cells (macrophages) play a critical role in combating RV loads and replication due to enhanced apoptosis of infected cells, IgE production, and concurrent exacerbation of Th2 responses in allergen-RV-induced asthma by inhibiting the production of CCL17, eotaxin-1, and eotaxin-2 (6081). Basically, type I IFNs (IFN-α/β) suppress Th2 cytokines via inhibition of IL 5 synthesis by CD4+ T cells, and interfere with the infiltration of eosinophils and CD4+ T cells into RV-infected areas (9). Despite the different mechanisms involved, type I and III IFNs share a similar role in the innate response against viral infections (79). A guinea pig model has been used to demonstrate the inhibitory role of type I IFN against the development of eosinophilic asthma after repeated OVA challenges (82). Similar mechanisms in human RV-infected lung epithelium treated with type I (IFN-β) and type III IFN-λ1 reduced the viral load and limited the viral replication (83).
Type II IFN
Type II IFN (IFN-γ) mimics a wide range of immunomodulatory activities compared with type I IFNs, with minimal antiviral innate response. IFN-γ upregulates the expression of MHC class I and II on the innate cells, particularly macrophages, and neutralizes the antigens presented by mast cells, the critical mediators of allergic airway inflammation. Moreover, IFN-γ may downregulate CCR3 expression (via eotaxin-1 and -2 receptors) and CXCL9 during induction, and the progenitor hematopoietic cells are regressed during the differentiation and activation of eosinophils (84). Several in vivo murine models also echoed the regulatory roles of allergen-induced asthma pathogenesis, and IFN-γ-receptor-ablated (GRKO) mice showed inhibition of prolonged eosinophilia via recruitment of CD4+ T cells and IL-5 induction (85). Besides, IFN-γ plays an important role in iNOS synthesis, resulting in IgE-mediated degranulation of mast cells, and inhibition of hyperplasia and hypertrophy of airway smooth muscle cells (86). Due to the anti-IgE class switching ability of IFN-γ, neutralization of inhaled allergen via FcγR-mediated endocytosis may epitomize the allergen capture mechanism of macrophages, and subsequent apoptosis of T cells and eosinophils via influx of proapoptotic proteins, caspases and Bax (87). Thus, the compelling inhibitory role of IFN-γ in allergic asthma demonstrates a definitive therapeutic approach, suggesting the need for further mechanistic studies using both in vivo and ex vivo models.
Type III IFNs
The innate roles of IFN-λ in RV-induced airway inflammation are mediated via a regulatory response in severe and exacerbated asthma (10). Type III IFNs (IFN-λ 1 and IFN-λ 2/3) display similar features, and share analogous roles in innate immunity after RV infection. RSV infection induces type III IFNs which activates Jak-Stat pathways to exhibit antiviral effect through ISG gene expression (88). Upon viral infection, TLR3, TLR4, TLR7, TLR8, and TLR9 mediate IFN-λ induction, and TLR3-mediated signaling is associated with RV-induced asthma severity and concurrent deficiency of IFN-λ (89). The ex vivo RV infection of isolated human bronchial epithelial cells and BAL cells derived from asthmatic subjects result in impaired IFN-λ synthesis (90). Moreover, the beneficial effect of IFN-λ in allergic asthma is clearly demonstrated using a murine model, where IL-28Rα−/− mice develop enhanced airway inflammation, increased mucus production, and a Th2-Th17-dependent cytokine response along with a reduction in IFN-γ and IL-12-expressing Th1 cells (91). Therefore, IFN-λ, despite lacking the multi-dimensional activity of type I IFNs, represents a potential candidate in combating RV-induced asthma exacerbation. Additional immunomodulatory mechanisms of type III IFNs should be investigated in different in vivo models.
Role of adaptive immunity in asthma exacerbation caused by respiratory viral infection
Cell-mediated immunity represents a comprehensive and more refined parameter for the recognition of self- and non-self-antigens, allergens and pathogens via regulation of interplay between APCs and lymphocytes (T and B cells). Thus, the adaptive immune response enhances antigen-specific immunological mechanisms to generate effector memory T cells and lead to concurrent immune homeostasis in the host.
B cells
The B cells have been the focus of attention following the detection of a series of B-cell-mediated immune responses in asthma, multiple sclerosis, and autoimmune diseases (92). In allergic inflammation or respiratory virus-induced asthma, B cells induce Ig synthesis effectively to overwhelm asthma progression. Isotype switching of B cells to allergen-specific IgE synthesis is highly dependent on IL-4 and IL-13 (via interaction with BCR), and provides the critical triggers during airway inflammation (93). The allergen-specific IgE interacts with low-affinity FcɛRII (CD23) receptor on B cells, and enhances both allergen-specific B and T cell-mediated recall in inflamed lungs (94). A cockroach allergen-induced allergy model showed a B cell-dependent prolongation of chronic pathogenesis mediated via Th2 phenotypes (95). The effector function of B cells is attributed to Be1 (B cells primed by Th1 cells) and Be2 (B cells primed with Th2 cells) cells, resulting in the differentiation of Th0 cells to IFN-γ-secreting Th1 and IL-4-dependent Th2 cells, respectively (96). Moreover, B lymphocytes induced T-cell-independent AHR and airway inflammation. The regulatory B cells manifested reversible inhibitory effects against allergic asthma via CD4+FoxP3+Treg cells in murine models (97).
T cells
The role of T cells in allergic airways and asthma exacerbation is mediated via complex mechanisms due to the different patterns of T cell subsets and effector functions involved in airway inflammation, remodeling, mucous cell metaplasia, intense and airway reactivity (AR), and less in prognosis. Notably, the function of CD4+ T cells is well known in allergic asthma based on their cytokine profiles; however, the CD8+ T cells also display an important protective or suppressive role in lungs. However, the functional role (regulatory/NK/proinflammatory) of CD8+ T cells needs to be characterized adequately. In inhaled allergic asthma, the CD8+ T cells exhibit proinflammatory, protective, and regulatory functions (83). After the onset of allergic inflammation or exacerbation with respiratory viruses, the CD4+ T helper (Th) cells display an adaptive immune response, and differentiate into effector T cell subsets secreting various cytokines (Fig. 2).
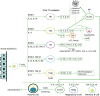 | Figure 2Contribution of CD4+ Th cells to airway inflammation. APCs including DCs process antigen (allergens/viruses) and present them on naive T cells, which subsequently induces the differentiation of naïve T cells into Th1, Th2, Th17, and Tregs cells, depending on the microenvironment orchestrated by the cytokine milieu. Following exposure to allergens, the Th2 response facilitates the accumulation of eosinophils in the airway via production of IL-4, IL-5, IL-9, and IL-13, whereas Th1 response is linked to mixed (eosinophil-neutrophil) inflammation via induction of IFN-γ, IL-8, TNF-α. Th17 cells mediate neutrophilic inflammation that is frequently observed in severe refractory asthma. CD4+Foxp3+ Tregs secreting IL-10 and TGF-β coordinate the tolerance to inflammation and balance of Th1/Th2 response.
|
In asthma pathogenesis, a robust and complex mechanism of differentiation and initiation of Th1, Th2, and Th17 cytokines or even Th9 response has been established using in vivo and in vitro models (17385860). In airway inflammation and allergic asthma, Th1 and Th2 responses are highly restricted via activation of transcription factors T-bet and GATA3, respectively (98) with subsequent cytokine production (99). The canonical Th2 response is mainly characterized by the production of IL-4, IL-5, and IL-13 (99); however, the IFN-γ-restricted Th1 response is proved to be amalgamated with Th2 cytokines (IL-4, IL-5, and IL-13), in chronic allergic responses (100). The exact mechanism underlying the differentiation of Th1 or Th2 response in atopic asthma or respiratory virus-induced exacerbation remains controversial, despite the balancing mechanisms of pro-inflammatory and suppressive Tregs (100). Moreover, Tregs provide insight into T-cell programming and flexibility during the activation of transcription factors to augment or repress cytokine gene expression during a stable Th cell response.
The most unpredictable (Th1 or Th17) features of severe asthma relate to the Th17 cell repertoire induced by IL-6, IL-22, and TGF-β mediated via expression of the transcription factor ROR-γt in several in vivo and ex vivo models of Th17 cells secreting IL-17A and IL-17F, and IL-25 (101). Recently, the Th17 cells were identified as prominent immune cells in airway inflammation of tissues during severe and chronic allergic asthma associated with steroid resistance. The most prominent and critical cytokine IL-17A binding to the IL-17R complex in the airway epithelium may act via increased NF-κB activation and induction of neutrophil chemokine, CXCL8 (102). The increased interest in the role of Th17 cells in severe asthma has been driven by their simultaneous appearance in classical Th2-mediated inflammation. Therefore, the Th17 immune responses can be orchestrated either alone or depending on Th1 or Th2 features, and also reciprocally with Th2 response (103). In addition to these unpredictable phenomena, IL-17A acts as a negative regulator of asthma by decreasing eosinophilic infiltration into the airways, AR, and expression of CCL5, CCL11, and CCL17 (104), and IL-17A neutralization exaggerated Th2-mediated eosinophilic response (105). Therefore, several in vivo and ex vivo models of allergic airway inflammation demonstrated the varying and conflicting roles of IL-17A and Th17 cells. Conversely, a murine model reinforced the synergistic signaling pattern of IL-23, and Th17 cells leading to increased eosinophilic Th2 response (106). Therefore, the precise and possible Th17-mediated mechanisms involved in allergic inflammation and RV-induced asthma exacerbation, are unknown.
The features of Th17 cell-dependent asthma triggered a new debate following the possible cross-talk between IL-17A and IL-22. Displaying both pro- and anti-inflammatory features, IL-22 is secreted from Th1, Th22, and Th17 cells with dominant IL-6, and TNF-α-dependent traits (107). A few reports suggested that CD4+ CCR6+ Th17 cells manifested the IL-22-secreting cellular phenotypes; however, lack of IL-22 resulted in a decreased Th2 response (107). Other studies reported contrasting asthmatic features associated with the induction of IL-17E, and IL-13 upon IL-22 neutralization in OVA-challenged asthma (108). Taken together, due to the pro- and anti-inflammatory roles of IL-22, several studies postulate a distinct role of CD4+Th22 cells in asthma progression.
Currently, the canonical asthma signatures have been used to address new concepts involving the critical differentiation of IL-9-dependent CD4+Th9 cells from Th2 cells (109). Previously, Th9 cells were treated as classical Th2 cells, due to their capacity to induce IL-9-dependent mast cell differentiation, goblet cell metaplasia, and IL-13 production (110). A few ex vivo models demonstrated Th9 cellular characterization upon IL-4, and TGF-β priming (109). Thus, in general, the CD4+Th9 cells induce Th2-like asthma features characterized by bronchial hyper-reactivity, mucus production, and airway eosinophilia following allergen exposure. However, the roles of Th2 and Th9 cells in asthma pathophysiology have been demonstrated adequately using eosinophil-deficient mice models (111). Therefore, an investigation into the Th9 cellular migration in asthmatic lungs is needed to elucidate the mechanism distinct from the Th2 response.
Allergic asthma or virus-induced severe airway inflammation resulting in Th1, Th2, Th17, Th22, Th9 or mixed inflammation can be controlled by Tregs to maintain airway tolerance against inhaled antigens (112). Usually, anti-inflammatory IL-10 plays a vital role in Treg function. Moreover, TGF-β also plays a bystander role in immune suppression via complex pathways to induce airway remodeling, and maintenance of immune homeostasis (113). Both thymic-derived and virus-induced CD4+FoxP3+Treg cells (iTregs/nTregs) disrupt the regulatory mechanism by inhibiting the effector T cell function, and the immunomodulatory role of subsequent APCs in airways, via IL-10 and TGF-β (114). Moreover, upon adaptive transfer, the antigen-specific CD4+CD25+ Tregs may also initiate allergic inflammation and AHR suppression via IL-10-dependent mechanism (115), and CD4+CD25+ Tregs induced the expression of the chemokine receptors CCR4 and CCR8 (116). Differentiation of highly secretory IL-10-dependent Tregs via ICOS-ICOS ligand pathway in asthmatic subjects was triggered by higher doses of inhaled allergens following chronic exposure, suggesting that repeated stimulation of airway DCs may strongly elevate IL-10 levels (117118).
In our preceding studies using the Japanese encephalitis virus mice model, we reported the roles of CCR5 chemokine receptor in the recruitment of CD4+FoxP3+Tregs during CNS inflammation (119). Similarly, we also found a diminished population of CD4+FoxP3+Tregs in CCR5-deficient mice following asthma exacerbation induced via allergen-sensitized RV-1B (unpublished personal data). Therefore, the CCR5 chemokine receptor also plays a role in the initiation and maintenance of CD4+Tregs. Although CD4+ Tregs received extensive attention, studies elucidating the role of CD8+ T cells, NK, γδ T cells, B cells, mast cells, and various APC populations in asthma are needed.
CONCLUSION AND FUTURE PERSPECTIVES
In asthma pathogenesis, a robust and complex cytokine connection has been established using numerous in vivo and in vitro models, demonstrating the differentiation and initiation mechanisms mediated via Th1, Th2, and Th17 or even Th9 response. The specific mechanism underlying the differentiation of Th1 or Th2 response in atopic asthma or respiratory virus-induced exacerbation (Th17 or Th17/Th2 mixed response) is intriguing, as the balancing roles of pro-inflammatory and suppressive Tregs suggest new paradigms and mechanisms associated with asthma prognosis.
Respiratory viral infections, particularly those associated with RVs, are the foremost cause of morbidity and mortality in exacerbated asthma due to poor prognosis, and resistance to corticosteroids. Patients diagnosed with allergic asthma are highly susceptible to respiratory virus and exhibit severe non-canonical asthma pathogenesis. Impaired antiviral immune responses are associated with severe disease burden, economic loss, disease severity and even mortality. Moreover, the specific anti-viral strategies are still under development in the absence of clear-cut efforts to prevent or combat virus-initiated asthma exacerbations. In addition to traditional ICS, the introduction of IL-4Rα is effective in Th2 asthma response by inhibiting the expression of IL-4 and IL-13. Moreover, mepolizumab, reslizumab, benralizumab, and dupilumab are also effective against classical asthma; however, they are associated with a poor response against virus-induced asthma characterized by a non-Th2 immune response. Different disease models enhance our understanding of the molecular epidemiology and mechanisms underlying disease progression. They provide opportunities for the development of antiviral therapies against specific targets for effective management and prevention. Hence, to delineate the virus-induced mechanisms exacerbating asthma for the development of appropriate antiviral therapeutics, extensive and sophisticated efforts are needed using effective in vivo and ex vivo models.
 XML Download
XML Download