

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Lead is a nonessential, toxic environmental contaminant and lead exposure and accumulation triggers various acute and chronic effects in humans [12]. Lead poisoning is associated with the most severe consequences during brain development, including differentiation, intense cellular proliferation, and synaptogenesis [3]. Acute exposure to lead during prenatal and postnatal periods is associated with psychosis, intellectual deficits, and juvenile delinquency [456]. Moreover, lead exposure during early life affects embryonic development. It is associated with negative outcomes including reduced neurocognitive development [78]. These effects manifest as deficits in social interaction, working memory, and spatial learning [910]. In addition, lead exposure impairs cognitive function and induces chronic changes in hippocampal and hypothalamic neurogenesis [11]. It also suppresses neuronal differentiation in central nervous system, inhibits longterm potentiation, inhibits the secretion of neurotransmitters, and interferes with calcium signaling [12]. All these processes affected by lead exposure are essential for neuronal function, synaptic plasticity, transmission of neurite growth, synaptogenesis, and axonal transport, which are required to maintain an intact microtubule structure [13]. Prenatal lead exposure affects embryonic development. It is also associated with negative outcomes in humans from birth to adulthood [7]. In addition, exposure to low levels of lead during early brain development impairs cognitive functions in children [14]. It also affects various cognitive domains, including attention, executive function, and social behavior [15]. Prepulse inhibition (PPI) is an operational parameter in sensorimotor gating. It has been hypothesized that PPI reflects the ability to filter extraneous interoceptive and exteroceptive stimuli [1617]. Reduced PPI has been observed in patients with schizophrenia and related disorders [1617]. It has been hypothesized that reduced PPI leads to impaired cognition reflecting gating of sensory input to the brain [18]. Several animal studies have demonstrated that PPI deficits can be induced by administration of dopamine agonists [19], NMDA antagonists [20], and exposure to low levels of lead [14]. Clozapine is an atypical antipsychotic agent and a synthetic dibenzodiazepine derivative, which inhibits several neurotransmitter receptors in the brain [21]. Moreover, clozapine was related to sensorimotor gating function that was restored in PPI and the mean% PPI scores after clozapine treatment of the maternal immune activation-induced schizophrenia animal model [22].
However, the effects of clozapine on lead-induced psychiatric disorders in an animal model of neurodegeneration in early life have yet to be reported. Therefore, the present study was designed to further characterize the pharmacological mechanisms of clozapine in psychiatric disorders in an animal model of prenatal lead exposure. Specifically, the present study determined the effect of clozapine on PPI modulation in startle, open-field, and social interaction tests using an animal model with chronic lowlevel lead exposure via oral route. Subsequently, we measured the protein exposure related to neurodevelopment in the hippocampus area via western blot and immunohistochemical analyses.
Schizophrenia is a neurodevelopmental disorder with a complex etiology comprising both genetic and environmental factors. Prenatal stress or maternal exposure induces cognitive deficits in offspring [23]. The cognitive deficits in schizophrenia are induced by disrupting the hippocampal anatomy and the function of hypothalamic-pituitary-adrenal axis [2425]. Dihydropyrimidinaselike 2 (Dpysl2) is a neurodevelopmental protein that regulates axonal outgrowth via promotion of microtubule, vesicle trafficking, and synaptic physiology in the developing brain [232425]. Moreover, Dpysl2 associated with disrupted in schizophrenia 1 (DISC1) interacts with many other proteins involved in synaptic function and neurodevelopment pathways related to schizophrenia and depression [2326]. Postsynaptic density protein 95 (PSD-95) is an essential component involved in glutamatergic transmission, synaptic plasticity, and dendritic spine morphogenesis during neurodevelopment. Interestingly, PSD-95 binds directly to DISC1 in high-risk schizophrenia and is responsible for synapse formation and microtubule network dynamics [2427].
METHODS
Animals and drugs
Sprague–Dawley female rats were purchased from Daehan Biolink, Ink (Eumsung, Korea) and reared at the animal facility of our institution. All animal procedures were performed in accordance with the guidelines for the care and use of laboratory animals of the United States National Institutes of Health [28]. The study protocol was approved by the Institutional Animal Care and Use Committee (IACUC) in Soonchunhyang University (Korea, IACUC approval No. SCH16-0059). Rats were provided free access to food and water. Pregnant females were individually housed in maternity cages on Gestation day 1. Lead acetate solution (0.2%) was prepared in tap water. Glacial acetic acid (0.5 ml/L) was added to lead acetate solution to prevent precipitation. Rats in the test group were exposed to 0.2% lead acetate in drinking water throughout the gestation and lactation periods (a total of 42 days). No mortality of rat dams was observed at weaning [20]. Clozapine (Sigma-Aldrich Co., St. Louis, MO, USA) was dissolved in 0.1N HCl to a concentration of 5 mg/kg/day. The animals were orally treated with clozapine solution during stress for 21 days. All groups contained litters of 7 to 12 pups with an equal number of males and females. Extremely large or small litters were eliminated. Offspring were weaned 23 days after birth and housed in groups. Male offspring were selected and used for further experiments. A total of three experimental groups of adolescents were tested: 1) control group comprising offspring of normal mothers; 2) LEAD group including offspring of mothers subjected to lead exposure before parturition; 3) LEAD-CLZ group, with clozapine-treated offspring in the adolescence period. Each group comprised 13 male rats.
Prepulse inhibition (PPI)
An automated startle reflex system (SR Lab, San Diego Instruments, San Diego, CA, USA) was used to measure PPI. The system consisted of a startle chamber housed in a sound attenuated isolation cabinet equipped with an internal fan and light. A cylindrical, transparent, acrylic holding apparatus resting on a fourpegged platform within the isolation chamber was used to hold each subject throughout the testing session (subject age, 57 days old). PPI is defined as the percent decrease in startle amplitude as a function of the magnitude of prepulse stimulus using the following formula: percent decrease = 100 − {[(startle response for prepulse + pulse trial) / (startle response for pulse alone trial)] × 100}. Mean % PPI was calculated based on the total prepulse test levels at 3, 6, and 12 dB above background [31].
Open field test (OFT)
OFT was used to assess the exploratory activity and reactivity to a novel environment. Subjects were removed from their home cages on test day (subject age, 58 days) and placed individually in an open-field start box (15 × 15 × 20 cm) for 5 min. The apparatus was constructed with black Polygal (77 × 77 × 25 cm). No background noise was detected. The experimenter exited the room and the subjects behavior was recorded for 20 min [293031].
Social interaction test (SIT)
SIT was adapted from previous studies and conducted in a black Polygal chamber (77 × 77 × 25 cm) (subject age, 59 days). The room in which the chamber was located was darkened during testing. The chamber was illuminated by a single 25 W red light bulb placed ~100 cm above the base of the chamber. Social interaction partners were same×sex siblings that resided in the same cage after weaning. They had approximately equal body weights. Each session lasted 20 min [3031].
Western blot
Hippocampus tissues were lysed (N = 5–6 animals per group) in RIPA buffer (EBA-1149; Elpis-Biotech, Inc., Daejeon, Korea) containing protease inhibitors and centrifuged at 14,000 rpm for 10 min at 4℃. To detect Dpysl2 and neurofilament protein M, 200 µg of lysed protein was subjected to 10 and 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to PVDF membranes (Millipore, Milford, MA, USA). After blocking with 5% skim milk, the membranes were incubated with anti-Dpysl2 (1:1,000; Cell Signaling Technology, Danvers, MA, USA), neurofilament M (Nefm; 1:1,000; Cell Signaling Technology), PSD95 (Dlg4; 1:1,000; Cell Signaling Technology), and β-tubulin (Tubb; 1:3,000; Thermo Fisher Scientific, Inc., Rockford, IL, USA) antibodies overnight at 4℃ followed by probing with peroxidase-conjugated secondary anti-mouse (1:10,000; Sigma-Aldrich Co.) and secondary anti-rabbit (1:5,000; Abfrontier, Seoul, Korea) antibodies for 1 h at room temperature. Immunoreactive bands were detected using an enhanced chemiluminescence kit (ELPIS-Biotech Inc., Daejeon, Korea).
Immunohistochemistry
Rats were deeply anesthetized with ethyl ether and perfused with 4% paraformaldehyde (N = 5–6 animals/each group). The fixed brains were removed, frozen, and cut into 30-µm thick sections for anatomical imaging of gene expression (6.24 mm interaural, −2.76 mm bregma, section 10 of the Rat Brain Library). The expression levels were averaged to CA3 area of hippocampus on the same side of the brain. Frozen sections from the hippocampus were blocked with normal horse serum, incubated with anti-Dpysl2 (1:1,000; Atlas Antibodies AB, Stockholm, Sweden), Nefm (1:100; Cell Signaling Technology), and NeuN (1:100; Millipore) followed by incubation with Cy3-conjugated anti-rabbit and mouse secondary antibodies (1:500 and 1:800 Jackson ImmunoResearch Laboratories, Inc., West Grove, PA, USA). Fluorescent images were captured using a confocal laser-scanning microscope (FV10-ASW; Olympus, Tokyo, Japan). Images were quantified with ImageJ software using a protocol described previously with slight modifications [3132].
Statistical analysis
All data are expressed as mean ± standard error of mean. They were compared using Student's t-test and/or ANOVA. If results of ANOVA indicated significant differences, a post-hoc analysis was performed with Tukey's honest significant difference (HSD) test. All statistical analyses were performed using IBM SPSS Statistics ver. 20 software (IBM Co., Armonk, NY, USA). Statistical significance was considered at p < 0.05.
RESULTS
Prepulse inhibition
Lead exposure during prenatal and lactation periods significantly affected sensorimotor gating. Results revealed significant differences in PPI levels among control, lead-exposed, and clozapine-treated groups. PPI levels at each stimulus level (3, 6, and 12 dB above background) in the lead-exposed group were not significantly changed compared to those in the control group (p > 0.05, Fig. 1A). However, the mean % PPI at total prepulse stimulus level was significantly different between control and lead-exposed groups (p < 0.05, Fig. 1B). Clozapine treatment significantly increased the mean % PPI level (p < 0.05, Fig. 1B).
Social interaction test (SIT)
All aggressive behavior scores during SIT were significantly increased in the offspring exposed to lead compared to those in the control group (p < 0.05, Table 1). Non-aggressive behavior scores of the lead-exposed offspring were decreased compared to those of the control group. Lead exposure also induced hyperactivity or excited states. The increased scores of aggressive behavior in the lead-exposed offspring were decreased to normal levels after clozapine treatment (p < 0.05, Table 1), but not at each prepulse stimulus level (p > 0.05, Fig. 1B).
Open-field test
The lead-exposed group significantly changed the number of central entries, line crossings, number and duration of rear, duration of cage sniff, and the degree of immobility compared to the control group. These scores were restored to near-normal levels after clozapine treatment (p < 0.05; Table 2). However, the frequency of immobility behaviors was increased in the leadexposed group, but reduced to control levels after clozapine treatment (p < 0.05, Table 2).
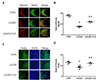
Western blot and immunohistochemistry
To investigate lead-induced changes in the expression of neurodevelopmental proteins such as Dpysl2, Dlg4, and Nefm, western blotting (Fig. 2) and immunohistochemical analyses (Fig. 3) of hippocampus areas from control, lead-exposed, and clozapinetreated offspring were performed. Western blot revealed that the expression of these proteins in the hippocampus of lead-exposed group was significantly decreased compared to those in the control (p < 0.05, Fig. 2B–D). These changes were restored by clozapine treatment (p < 0.05, Fig. 2B–D). Dpysl2 and Nefm were differentially expressed in immunofluorescence-stained brain images among control, lead-exposed, and clozapine-treated groups (Fig. 3A, C). The expression of Dpysl2 and Nefm proteins in the hippocampus of lead-exposed group was significantly decreased compared to that of the control (Fig. 3A, C). These changes were restored by clozapine treatment (Fig. 3A, C). The immunohistochemical staining intensity also varied significantly among the three groups (p < 0.05, Fig. 3B, D).
DISCUSSION
In this study, impaired social interaction was observed in rat offspring exposed to lead. In social interaction test, the diminution of social interaction behavior reflected increased anxiety, hyperactivity, and aggressive behaviors in lead-exposed animal models. Lead-induced increases in aggressive behavior and hyperactivity were attenuated by clozapine treatment in the present study (Table 1). Additionally, a few behavioral patterns in open field test were reversed by clozapine treatment (Table 2). These results are consistent with previous studies showing that lead exposure may cause hyperactivity and deficits in learning and memory both in experimental animals and humans [31333435]. In PPI test, lead exposure failed to significantly affect the startle amplitude in a no-PPI trial in a previous study, but not startle amplitude at 120 dB [36]. Reduction in PPI of acoustic startle associated with chronic exposure to low-level oral lead might be attributed to lead-induced alteration in glutamate neurotransmission [37]. Clozapine appears to increase low PPI in normal subjects, consistent with a few studies investigating sustained treatment with atypical antipsychotics and schizophrenia [38]. Prenatal viral mimic-induced PPI disruption was normalized by pre-adolescent clozapine treatment. However, clozapine failed to significantly reduce % PPI in adulthood, although it significantly reduced the mean % PPI [38]. Our results also demonstrated that clozapine treatment significantly reduced the mean % PPI level (p < 0.05, Fig. 1B) without significantly affecting acute startle response (p > 0.05, Fig. 1A). These results suggested that our animal model was affected by low-level lead exposure probably due to inhibition of sensorimotor gating function caused by altered glutamate neurotransmission induced by NMDA antagonist. Several studies suggested that lead-induced neurotoxicity decreased the expression of NMDAR subtypes, and reduced the calcium-dependent synaptic transmission [3940].
We evaluated whether maternal lead exposure affected the expression of PSD95, Dpysl2, Lasp1, and Nefm. PSD95 is essential for post-synaptic connectivity, downstream signaling cascade, neuronal maturation, and plasticity [4142]. The reduction in these proteins was expected to affect PSD95-NMDA receptor complex formation in postsynaptic membrane sites and related downstream signals, and alter brain development and cognition in lead-exposed dams. This speculation was consistent, at least partially, with previous reports showing that lead exposure during synaptogenesis led to NMDA receptor inhibition [4142]. Furthermore, lead impaired slow axonal transport in rats with hyperphosphorylated neurofilaments, reducing the mobility of neurofilaments that are not extensively phosphorylated [43]. Our previous study has shown that NMDA antagonism may downregulate the expression of proteins such as Lasp1 in a schizophrenia-like animal model [44]. Several studies have reported that a schizophrenia-like model induced by prenatal stress showed altered expression of NMDA receptor as one of the glutamate receptors involved in neurodegeneration, learning, and memory [4546]. Our recent study using another schizophrenia-like animal model showed that the expression of Dpysl2 was changed in prefrontal cortex and hippocampus area [23]. Therefore, proteins associated with brain development and cognition might be downregulated due to NMDA receptor inhibition caused by lead exposure. In the present study, our results indicated that decreased levels of brain development and cognition-related proteins can be restored by clozapine treatment. Clozapine is an effective partial agonist of muscarinic M1 receptors and an agonist of dopamine D1 receptors resulting in enhanced NMDA-R function [47]. Hence, clozapine may increase the NMDA-R transmission, explaining the unique efficacy of this agent. A recent study reported that the expression of SNPA-25, a synaptic gene, was influenced by clozapine treatment in male rats [47]. Although our data provided direct evidence suggesting that lead exposure decreased the levels of brain development and cognition-related proteins, the mechanism of decrease and recovery by clozapine treatment remains unclear. Therefore, further studies are needed to evaluate the role of clozapine for the treatment of lead exposure-induced neurodevelopmental psychiatric disorders such as autism and schizophrenia. In addition, cellular and animal models exposed to various levels of lead exposure are needed to determine the mechanism underlying the effect of clozapine.
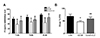



 XML Download
XML Download