

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Acute myeloid leukemia (AML) is a type of blood malignancy which involves cells that differentiate into white blood cells. AML starts in the bone marrow (BM). It comprises a heterogeneous group of disorders characterized by the rapid expansion of immature myeloid cells (blasts) in the BM. The inability of current therapies to eradicate blasts and chemotherapy refractoriness are the major causes underlying AML progression/relapse. The high rate of mortality due to AML reinforces the need for a greater understanding of the leukemic BM microenvironment and alternative methods for treating this disorder. Stem cell therapy is one of the best candidates to treat such hematological malignancies. There are different types of stem cells, including embryonic stem cells (ESCs), fetal stem cells, mesenchymal stem cells (MSCs), and hematopoietic stem cells (HSCs) that continuously replenish certain tissues. Among the different types of stem cells, MSCs have been specifically considered for clinical applications. Since the discovery of MSCs, various studies were performed to understand their physiology, function, and behavior [1]. Therefore, it is generally reported that MSCs have multi-lineage potential and are capable of differentiating into various types of cells [2]. The multi-lineage capacity of MSCs makes them promising therapeutic targets and one of the most indispensable sources of cell therapy and regenerative medicine resources. Despite comprehensive research performed over the past 10 years, it is still unknown whether MSCs have tumor-promoting or tumor-suppressing effects. Research is required before MSCs can be used for the treatment of hematologic malignancies. In this review, we give an overview of studies regarding the use of MSCs in the treatment of AML as an example of an evolving model of myeloid malignancies (Table 1).
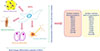
CHARACTERIZATION AND APPLICATION OF MSCS
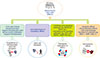
The pioneering work of Caplan in 1991 introduced the terms stroma and MSCs to the scientific community and indicated that MSCs are able to differentiate into the adipocyte cell lineage [3]. MSCs, as well as other types of adult stem cells, are characterized by self-renewal capability, clonogenic efficiency, and multi-lineage differentiation capacity. In general, MSCs are isolated by their capacity to adhere to culture-dish plastic surfaces. Cells can be expanded in culture plates and immunologically characterized by a specific panel of markers. Because of the lack of unique and definitive cellular markers, the characterization of MSCs remains difficult. For this reason, the International Society for Cellular Therapy suggested three minimal criteria for the characterization of MSCs: (a) plastic adherence, (b) expression of markers related to mesenchymal cells such as CD73, CD90, and CD105, and lack of hematopoietic-related cells such as CD34, CD45, CD11b or CD14, CD19 or CD79α, and HLA-DR expression, and (c) their tri-lineage differentiation potential into adipocytes, osteoblasts, and chondrocytes (Fig. 1) [4]. MSCs, which are present in adult tissues and organs such as adipose tissue, liver, kidney, heart, placenta, amniotic fluid, amnion, and BM, among others, are undifferentiated cells that have the capacity to differentiate into a broad range of different cell types, including adipocytes, neuron-like cells, osteocytes, chondrocytes, and other connective tissues [56789]. Earlier, it was demonstrated that MSCs can only differentiate into mature cells of the same organ, but recent findings have shown that these cells can also differentiate into other cell types, and even into the cells making up endoderm, mesoderm, and ectoderm [10]. Also, due to their plasticity, self-renewal, and relatively non-immunogenic properties, MSCs are potentially considered for regeneration, transplantation, and treatment of certain diseases such as ischemia, multiple sclerosis, cartilage and bone pathologies, cardiac events, auto-immune disorders, cancer, genetic diseases, and blood malignancies [1112]. Of the forementioned diseases, hematological abnormality and blood malignancy have gained more attention for cell transplantation with MSCs (Fig. 2) [13]. Some studies indicated that MSCs could release multiple angiogenic growth factors and cytokines/chemokines. These observations suggest that MSCs may have potential as a useful cell source for therapeutic angiogenesis [14]. Following these findings, Fernandez-Garcia et al. (2015) reported that adipose tissue derived-MSCs (ADSCs) improve the homing of donor HSCs and progenitor cells into recipient BM, facilitating the stable reconstitution of transplanted recipients with infused hematopoietic grafts [15]. These results open new perspectives for the application of ADSCs in HSCs therapy. Furthermore, the therapeutic potential of MSCs in veterinary medicine has been demonstrated since 2003 and MSC-based therapies have used in more than 500 dogs and 2,500 horses [16]. However, there are concerns regarding these cells and the risks linked to their therapeutic use are still unclear, particularly in the context of patients affected by pre-existing cancer [17]. It was reported that interactions between cancer cells and MSCs are of fundamental importance in stimulating both the development and invasiveness of tumors. As mentioned above, MSCs have specific features that make them candidates for cell therapy. One of the known roles of these cells was indicated by their use for the treatment of a hematological disorder [17].
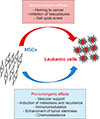
ROLE OF MSCS IN HEMATOLOGIC MALIGNANCY PROGRESSION
Cancer cells involve a set of abnormalities, including uncontrolled cell growth, cell invasion, genetic instability, and, finally, tumor development and metastasis. Therefore, a variety of promising new therapies for cancers, such as immunomodulation and cell therapy, are being developed. Ongoing studies propose that MSCs are good targets for cell therapy in a variety of cancers. The effects of MSCs on cancer cells are yet to be controversial [13]. Some studies indicated inhibitory effects, while others reported proliferative activity. For instance, in an in vitro study, it was shown that MSCs have tumoricidal effects on breast and lung cancer cell lines [18]. Furthermore, the co-culture of MSCs and melanoma cancer cell line cells revealed the promotion of cell proliferation [19]. It was also shown that MSCs cause tumor growth when injected into mice with prostate cancer [20]. It seems that MSCs, through signaling pathways, can suppress both proliferation and apoptosis of cancer cells [2122]. This dual role of MSCs can be described as a “double-edged sword”. Therefore, understanding MSCs' dual roles in tumor cell proliferation and apoptosis is required. Identifying the function of MSCs in hematologic malignancies such as lymphoma and leukemia may be applicable to hematologic cancer treatment. Regarding some studies involving hematologic malignancies, it has been shown that MSCs are capable of promoting or inhibiting tumor growth by suppressing apoptosis, or proliferation of tumor cells, respectively (Fig. 3) [1131823]. Although few investigations have reported that MSCs can directly promote the proliferation or apoptosis of malignant cells, the predominant hypothesis is that MSCs suppress both proliferation and apoptosis [2425]. Thus, the efficacy of the use of MSCs in treatment of hematologic malignancies is poorly understood, and the mechanisms underlying the pro-tumorigenic and anti-tumorigenic effects of these cells act are currently unclear [26]. Some studies suggest mechanisms that have inhibitory effects on hematologic malignancies. In general, the mechanisms mentioned include the possible use of MSCs as a delivery vehicle [27], inhibition of vascular growth [28], or to arrest the cell cycle [2629].
ANTI-TUMORIGENIC EFFECTS OF MSCS
Various studies indicated that MSCs can interfere hematologic malignancies via inhibiting the proliferation of tumor cells. Various sources of MSCs have been utilized for this purpose. These sources include BM, which was the first source of MSCs discovered for clinical applications; umbilical cord blood; and adipose tissue [30]. MSCs derived from these sources are known to have similar surface antigen expression phenotypes, and immunosuppressive properties [31]. Our experimental results showed that the antitumor effects of MSCs are not dependent on their tissue source and origin. Besides the cell source used, the number of MSCs and cancer cells seeded for co-culture is another important consideration. In other words, culture conditions, especially the concentration of MSCs, are known to significantly affect proliferation rate, morphology and secreted factors [32]. Moreover, it has been reported that antitumor effects in solid tumors are observed to associate with a higher number of MSCs [23]. This dependency is yet to be suggested for hematologic malignancies. There are many proposed mechanisms describing the effects of MSCs on cancer cells; however, the most commonly-accepted mechanism is that MSCs induce tumor cell cycle arrest. Most studies related to effect of MSCs on AML cells were carried out using U937, HL-60, and HL-60/VCR cell lines instead of primary cells. In this regard, Liang et al. (2008) reported that direct contact of U937, HL-60, and HL-60/VCR AML cells with human BM fibroblast stromal cells (HFCLs) causes inhibition of cell proliferation and induction of apoptosis. In their study, it was shown that upon co-culture with HFCLs, the percentage of AML cells in the G1 phase was higher and that of AML cells in the S phase cells was lower than those without HFCL cell-coculture [33]. In other words, Liang et al. (2008) suggested cell cycle G0/G1 blockage by transcriptional activation of specific genes [33]. In parallel, MSCs were found to inhibit the self-renewal ability of tumor cells. In this regard, Tian et al. (2010) demonstrated that umbilical cord (UC)-MSCs cause proliferative inhibition of HL-60 cells due to G0/G1 arrest. In addition, in this study, p38 mitogen- activated protein kinase (MAPK) was suggested as a potent suppressor of cell proliferation and tumorigenesis in this cell line [34]. In another investigation, Li et al. (2018) showed that UC-MSCs inhibited the proliferation of HL-60 and THP-1 cells as AML cell lines. Their results indicate varying effects of UC-MSCs on various types of AML cell lines associated with secreted cytokines and the expression of cytokine receptors on the cells [35]. They suggested different mechanisms, such as secretion of certain substances or paracrine signals, for the antitumor effects of UC-MSCs, besides cell cycle arrest, but the exact mechanism was not determined in their study.
FAVORABLE AND POSSIBLY-INVOLVED MECHANISMS OF MSCS FOR INHIBITION OF HEMATOLOGIC MALIGNANCY
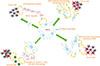
MSCs as delivery vehicles
As promising delivery vehicles, MSCs can be used for cancer cell therapy (Fig. 4) [3637]. These cells are easily accessible, quickly cultured in vitro, and transplanted [38]. Moreover, MSCs are known to possess hypo-immunogenic characteristics and can migrate to tumor sites [39]. It has been reported that the cytotoxic effects of MSCs may be helpful if they could identify tumor sites and migrate to it [40]. However, there have been a number of challenges in adapting the homing ability of MSCs for targeted delivery [41]. Furthermore, MSCs can also be used as gene therapy carriers [42]. In the same way, several studies have used MSCs to deliver genes and other factors, such as IL-12 [43], IL-24 [44], and IFN-γ [45] to tumor sites. Moreover, at the cellular level, MSCs exert most effects through extracellular vesicles (EVs), including microvesicles and exosomes [46]. These EVs are lipid membrane-bound vesicles secreted from MSCs. EVs contain a variety of molecules such as microRNAs, RNA, and proteins that have originated in MSCs, and these contents can be transferred to other cells, such as cancerous cells [47]. In one study by Hendijani et al. (2015), it was reported that MSC EVs produced an anti-proliferative effect on leukemic cells, and a cytotoxic effect in combination with doxorubicin, demonstrating an anti-leukemic potential of exosome-derived MSCs [48].
Inhibition of vascular growth
In recent studies, it was shown that tumor growth resulted from proangiogenic characteristics of MSCs. However, it has been documented that MSCs can impair vessel growth or angiogenesis under certain conditions. They can migrate to endothelial cell-derived-capillaries to produce reactive oxygen species (ROS) [49]. Following generation of ROS, apoptosis of endothelial cells and suppression of tumor growth take place [28]. The mechanisms involved relate to modulation of the vascular endothelial cadherin/β-catenin signaling pathway [50]. Moreover, previous studies reported that MSCs present in high numbers are potentially cytotoxic, and injection of MSCs into tumor sites may be an effective antiangiogenic treatment [49]. However, the inhibitory effect of MSCs on tumor growth has not been clearly indicated in hematologic cancers, but it may be important as these cancers are dependent on vascular support.
Cell cycle arrest
Cell cycle arrest is the most common fundamental process of tumor growth inhibition. Although cell cycle checkpoints and DNA repair processes seem to be linked to various cancers, mechanisms inducing cancer cell-cycle arrest by antitumor agents are yet to be identified. In other words, the effects of MSCs on leukemia, lymphoma and another blood malignancies are not well understood [51]. Some studies have shown high levels of cells arrested at G0/G1. In detail, Fonseka et al. (2012) indicated that UC-MSCs could inhibit the proliferation of K562 cells due to arrest in G0/G1 phase through IL-6 and IL-8 cytokine secretion [52]. In another study, Fathi et al. (2019) reported that BM-derived MSCs were attributed to a robust increase in the number of cells in G0/G1 phase, which implies cell arrest at G0/G1. This result agrees with earlier reports by other authors [5253]. Therefore, further research is needed to understand the mechanisms of tumor cell cycle arrest that consequently lead to the antitumor effects of MSCs in hematologic malignancies.
MSC behavior in myeloid malignancies
As mentioned above, MSCs exhibit immunoregulatory properties that influence both innate and adaptive immune responses [5455]. Moreover, it seems that MSCs inhibit erythropoiesis to favor myeloid differentiation via production of cytokines and growth factors, such as interleukin (IL)-6, which was shown to expand myeloid progenitors and block erythroid development [56]. In this context, elevated IL-6 levels have been correlated with adverse survival in patients with AML [57]. Another player engaged in the BM microenvironment (niche) regulation is the autonomic nervous system that accompanies marrow blood vessels through adrenergic fibers [58]. Deregulation of the interaction between adrenergic fibers and the MSC niche has been implicated in impaired hematopoiesis, which is a hallmark of several hematologic diseases [58]. As previously reported, as an evolving model of myeloid malignancies, AML-derived MSCs display enhanced supportive capacity for hematopoiesis by changing expression of cell surface molecules or CD markers, including CD44, CD49e, CD271 and CXCL12 [5960]. As confirmed for myelodysplastic syndrome, in AML, MSC-derived endothelial cell numbers are predominantly increased, especially in cases with rapidly proliferating disease, further suggesting MSC-derived cell implication in leukemic niche building. Moreover, it has been indicated that AML blasts can modulate endothelial cell expansion via the upregulation of E-selectin adhesion molecules and may then adhere to the stroma and be concealed in a silent status, eventually becoming chemo-resistant [61]. Recent evidence, reviewed by Huang et al. (2015), report that AML-derived MSCs from AML patients show similar CD90, CD73, CD44, and E-cadherin expression, but decreased monocyte chemoattractant protein-1 levels compared to MSCs from healthy donors [62]. Also, AML blast interactions with MSCs show that both cells release several cytokines and chemokines and, when co-cultured, normal MSCs had an anti-apoptotic and growth-enhancing effect on primary human AML cells, this was associated with upregulation of the mTOR signaling pathway [63]. Recently, it has been shown that different clinical/cytogenetic AML subgroups may show differences in MSC niches. In one study, Lopes et al. (2017) characterized and arrayed MSC cytokine expression in patients with de novo AML and secondary AML [AML with myelodysplasia related changes (MRC)]. They found that de novo AML-derived MSCs presented VEGFA, CXCL12, RPGE2, IDO, IL-1β, IL-6, and IL-32 at high levels and IL-10 in lower levels. However, AML-MRC-derived MSCs presented IL-6 at high levels [64].
MESENCHYMAL STEM CELLS: THERAPEUTIC CONCEPTS VIA TARGETING IMMUNE ESCAPE
Immune dysregulation of leukemic niches is an attractive approach for cellular therapies. Recently, an increasing number of reports have supported the use of immune checkpoint blockers as well as monoclonal antibody therapies engaging specific T cells in hematologic malignancies. Immune checkpoints are one of the protective mechanisms that are induced in activated T cells and which regulate T cell antigen responses. In other words, cancers can evade immune-mediated destruction by upregulation of certain molecules on the surface of T cells. Indeed, immune checkpoint blockers could enhance cytotoxicity of cytokine-induced killer cells against myeloid leukemic blasts [65]. Recently it was shown that vaccination with MSCs promotes apoptosis of tumor cells and inhibits proliferation by increasing MHC1 and heat shock protein (HSP) expression levels. In detail, the enhanced antitumor response of MSCs was strongly associated with higher expression levels of MHC class I molecules on dendritic cells (DCs) that made tumor cells more cross-presentable to host DCs to generate antitumor activity [66]. Another attractive perspective includes the optional transfer of gene-modified MSCs which secrete tumor-directed antibodies continuously into the body of the patient. As MSCs have less immunogenicity and tend to condense in the close neighborhood of the tumor, they can be used as a means for the targeted delivery of anticancer agents. Aliperta et al. (2015) reported that gene-modified MSCs are able to express a CD33-CD3 bispecific antibody and to interfere with efficient lysis of AML blasts by human T cells in AML patients [66]. With regard to antibody-derived agents, such as bispecific agents and antibody-drug conjugates, CD33 is a clinically validated target and was shown to be effective in AML treatment [66]. In addition, antibodies specific for CD123 are under evaluation [67]. Li et al. (2018) indicated that the anti-CD44 antibody A3D8 inhibits proliferation of HL-60 cells, a representative acute leukemia cell line [35]. The A3D8 treatment increased the percentage of cells in G0/G1 cell cycle phase [68]. However, other in vitro investigations reported that MSCs may escape this targeted therapy and that leukemic stem cells become less microenvironment-dependent in advanced-stage AML, so that targeting of CD44 may be less successful than expected. Other attractive therapeutic approach for myeloid disorders involve the use of allogeneic BM transplantation, chimeric antigen receptor T (CART) cells, and donor lymphocyte infusion (DLI) [69]. These approaches are presently aimed at targeting leukemic blasts, but the use of MSCs might be novel targets in the near future.
CONCLUSIONS
MSC-based therapeutic approaches have shown a wide range of outcomes, probably due to non-standardized experimental methods, heterogeneous characteristics of MSCs, and a lack of specific cell surface markers that are easily affected by the surrounding environment. The tumor-related effects of MSCs are still not well understood. Therefore, much more researches are necessary to develop MSCs as a cell-based therapy for cancer. Various studies have been carried out to investigate the effects of MSCs in tumorigenesis, but a single principle cannot explain the dual anti-tumorigenic and pro-tumorigenic roles of MSCs. It has been indicated that the antitumor effects of MSCs are principally a result of the suppressed proliferation of malignant cells via an arrest in the G0/G1 phase of the cell cycle [23]. In order to exploit this anti-tumorigenic feature of MSCs for clinical use in the future, more investigation is recommended.

 XML Download
XML Download