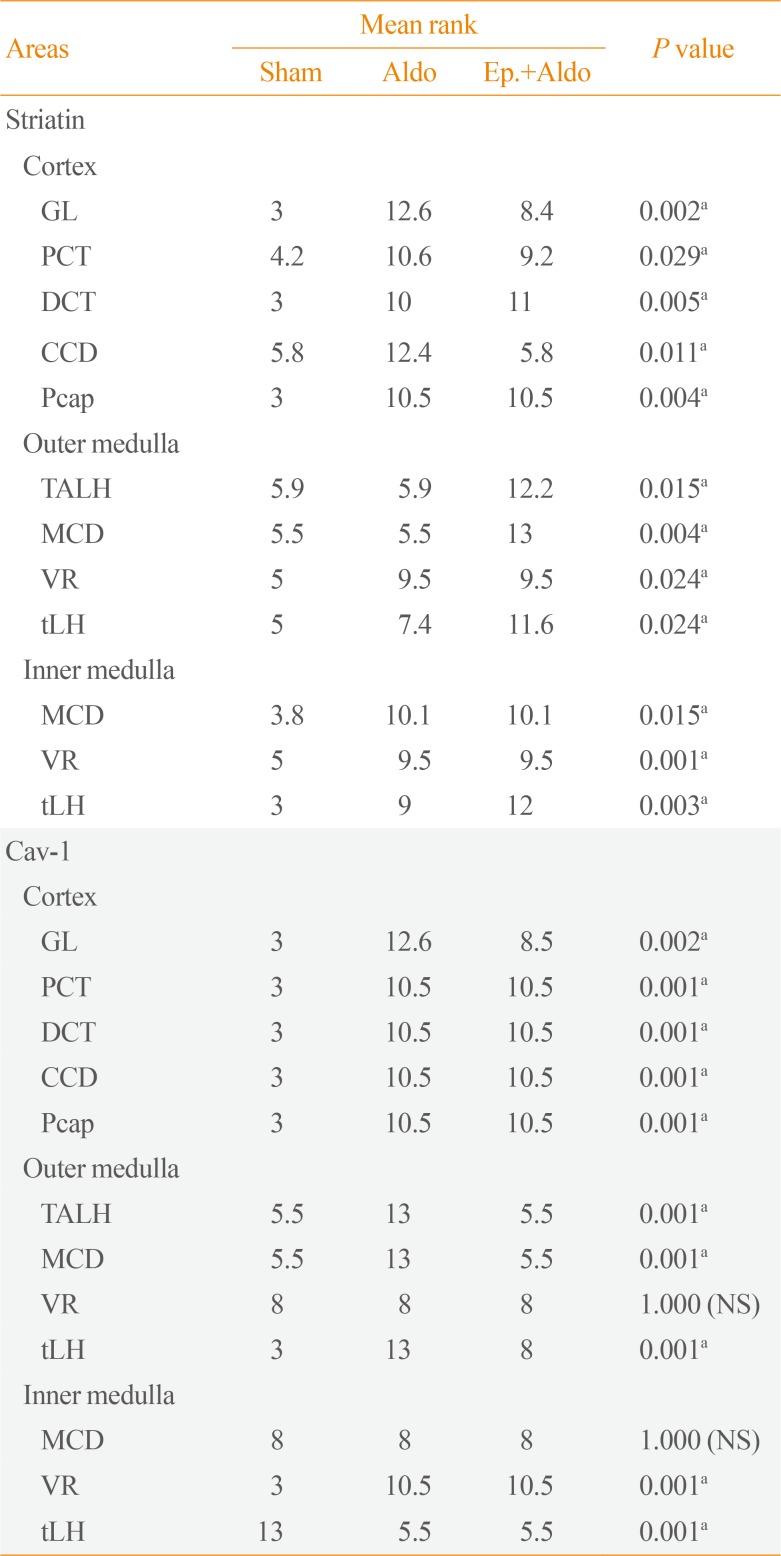
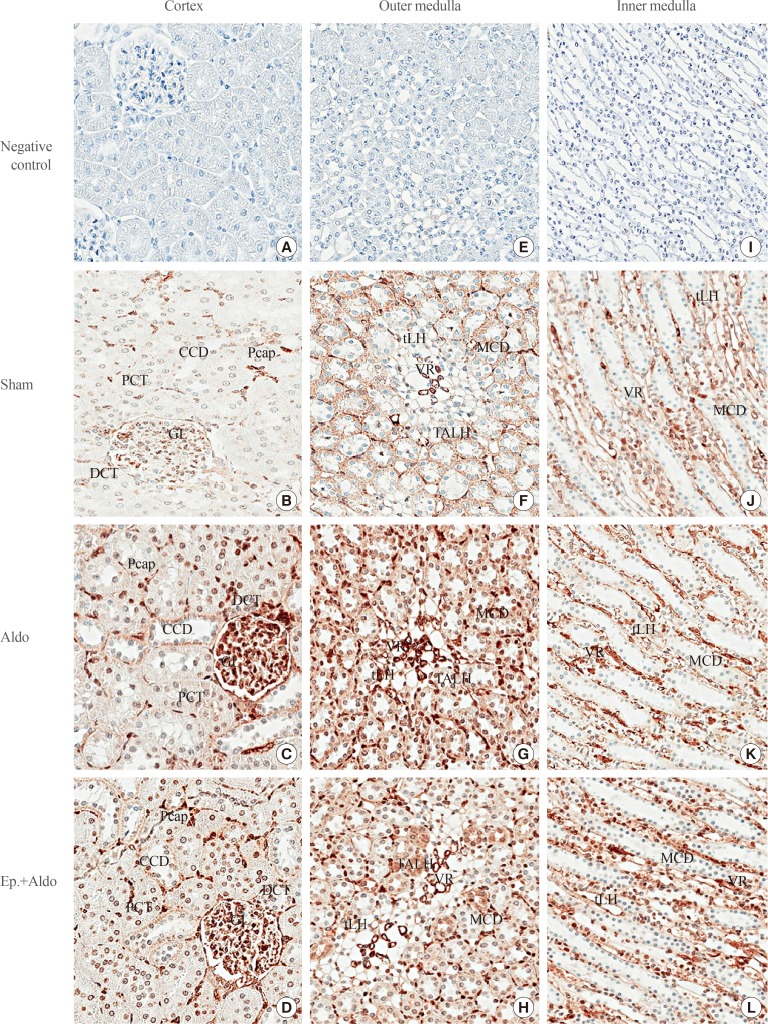
1. Hwang J, Pallas DC. STRIPAK complexes: structure, biological function, and involvement in human diseases. Int J Biochem Cell Biol. 2014; 47:118–148. PMID:
24333164.

2. Baudrand R, Pojoga LH, Romero JR, Williams GH. Aldosterone's mechanism of action: roles of lysine-specific demethylase 1, caveolin and striatin. Curr Opin Nephrol Hypertens. 2014; 23:32–37. PMID:
24275769.
3. Garza AE, Rariy CM, Sun B, Williams J, Lasky-Su J, Baudrand R, et al. Variants in striatin gene are associated with salt-sensitive blood pressure in mice and humans. Hypertension. 2015; 65:211–217. PMID:
25368024.
4. Williere Y, Borschewski A, Patzak A, Nikitina T, Dittmayer C, Daigeler AL, et al. Caveolin 1 promotes renal water and salt reabsorption. Sci Rep. 2018; 8:545. PMID:
29323234.
5. Bollag WB. Regulation of aldosterone synthesis and secretion. Compr Physiol. 2014; 4:1017–1055. PMID:
24944029.
6. Connell JM, Davies E. The new biology of aldosterone. J Endocrinol. 2005; 186:1–20. PMID:
16002531.
7. Grossmann C, Gekle M. New aspects of rapid aldosterone signaling. Mol Cell Endocrinol. 2009; 308:53–62. PMID:
19549592.
8. Falkenstein E, Wehling M. Nongenomically initiated steroid actions. Eur J Clin Invest. 2000; 30:51–54. PMID:
11281368.
9. Thomas W, Harvey BJ. Mechanisms underlying rapid aldosterone effects in the kidney. Annu Rev Physiol. 2011; 73:335–357. PMID:
20809792.
10. Dooley R, Harvey BJ, Thomas W. Non-genomic actions of aldosterone: from receptors and signals to membrane targets. Mol Cell Endocrinol. 2012; 350:223–234. PMID:
21801805.
11. Tsugita M, Iwasaki Y, Nishiyama M, Taguchi T, Shinahara M, Taniguchi Y, et al. Glucocorticoid receptor plays an indispensable role in mineralocorticoid receptor-dependent transcription in GR-deficient BE(2)C and T84 cells in vitro. Mol Cell Endocrinol. 2009; 302:18–25. PMID:
19146914.
12. Pojoga LH, Coutinho P, Rivera A, Yao TM, Maldonado ER, Youte R, et al. Activation of the mineralocorticoid receptor increases striatin levels. Am J Hypertens. 2012; 25:243–249. PMID:
22089104.
13. Coutinho P, Vega C, Pojoga LH, Rivera A, Prado GN, Yao TM, et al. Aldosterone's rapid, nongenomic effects are mediated by striatin: a modulator of aldosterone's effect on estrogen action. Endocrinology. 2014; 155:2233–2243. PMID:
24654783.
14. Pojoga LH, Romero JR, Yao TM, Loutraris P, Ricchiuti V, Coutinho P, et al. Caveolin-1 ablation reduces the adverse cardiovascular effects of N-omega-nitro-L-arginine methyl ester and angiotensin II. Endocrinology. 2010; 151:1236–1246. PMID:
20097717.
15. Igarashi J, Hashimoto T, Shoji K, Yoneda K, Tsukamoto I, Moriue T, et al. Dexamethasone induces caveolin-1 in vascular endothelial cells: implications for attenuated responses to VEGF. Am J Physiol Cell Physiol. 2013; 304:C790–C800. PMID:
23426970.
16. Sinphitukkul K, Eiam-Ong S, Manotham K, Eiam-Ong S. Nongenomic effects of aldosterone on renal protein expressions of pEGFR and pERK1/2 in rat kidney. Am J Nephrol. 2011; 33:111–120. PMID:
21196724.
17. Eiam-Ong S, Sinphitukkul K, Manotham K, Eiam-Ong S. Rapid nongenomic action of aldosterone on protein expressions of Hsp90 (α and β) and pc-Src in rat kidney. Biomed Res Int. 2013; 2013:346480. PMID:
23484111.
18. Eiam-Ong S, Sinphitukkul K, Manotham K, Eiam-Ong S. Rapid action of aldosterone on protein expressions of protein kinase C alpha and alpha1 sodium potassium adenosine triphosphatase in rat kidney. J Steroids Horm Sci. 2014; 5:125.
19. Eiam-Ong S, Chaipipat M, Manotham K, Eiam-Ong S. Rapid action of aldosterone on protein levels of sodium-hydrogen exchangers and protein kinase C beta isoforms in rat kidney. Int J Endocrinol. 2017; 2017:2975853. PMID:
29201052.
20. Chen YK, Chen CY, Hu HT, Hsueh YP. CTTNBP2, but not CTTNBP2NL, regulates dendritic spinogenesis and synaptic distribution of the striatin-PP2A complex. Mol Biol Cell. 2012; 23:4383–4392. PMID:
23015759.
21. Zhang J, Jiang Z, Bao C, Mei J, Zhu J. Cardiopulmonary bypass increases pulmonary microvascular permeability through the Src kinase pathway: involvement of caveolin-1 and vascular endothelial cadherin. Mol Med Rep. 2016; 13:2918–2924. PMID:
26847917.
22. Machado-Fiallo ED, Vega C, Ramos-Rivera A, Benabe-Carlo JA, Prado GN, Pojoga LH, et al. Striatin is a novel regulator of aldosterone-mediated endothelial cell activation. Blood. 2013; 122:3523.
23. Nagoshi T, Date T, Fujisaki M, Yoshino T, Sekiyama H, Ogawa K, et al. Biphasic action of aldosterone on Akt signaling in cardiomyocytes. Horm Metab Res. 2012; 44:931–937. PMID:
22763651.
24. Zheng S, Sun P, Liu H, Li R, Long L, Xu Y, et al. 17β-Estradiol upregulates striatin protein levels via Akt pathway in human umbilical vein endothelial cells. PLoS One. 2018; 13:e0202500. PMID:
30138337.
25. Zheng S, Chen X, Hong S, Long L, Xu Y, Simoncini T, et al. 17β-Estradiol inhibits vascular smooth muscle cell migration via up-regulation of striatin protein. Gynecol Endocrinol. 2015; 31:618–624. PMID:
26220767.
26. Luo X, Dan Wang, Luo X, Zhu X, Wang G, Ning Z, et al. Caveolin 1-related autophagy initiated by aldosterone-induced oxidation promotes liver sinusoidal endothelial cells defenestration. Redox Biol. 2017; 13:508–521. PMID:
28734243.
27. Braun S, Losel R, Wehling M, Boldyreff B. Aldosterone rapidly activates Src kinase in M-1 cells involving the mineralocorticoid receptor and HSP84. FEBS Lett. 2004; 570:69–72. PMID:
15251441.
28. Callera GE, Montezano AC, Yogi A, Tostes RC, He Y, Schiffrin EL, et al. c-Src-dependent nongenomic signaling responses to aldosterone are increased in vascular myocytes from spontaneously hypertensive rats. Hypertension. 2005; 46:1032–1038. PMID:
16157790.
29. Callera GE, Touyz RM, Tostes RC, Yogi A, He Y, Malkinson S, et al. Aldosterone activates vascular p38MAP kinase and NADPH oxidase via c-Src. Hypertension. 2005; 45:773–779. PMID:
15699470.
30. Li S, Seitz R, Lisanti MP. Phosphorylation of caveolin by src tyrosine kinases. The alpha-isoform of caveolin is selectively phosphorylated by v-Src in vivo. J Biol Chem. 1996; 271:3863–3868. PMID:
8632005.
31. Tourkina E, Gooz P, Pannu J, Bonner M, Scholz D, Hacker S, et al. Opposing effects of protein kinase Calpha and protein kinase Cepsilon on collagen expression by human lung fibroblasts are mediated via MEK/ERK and caveolin-1 signaling. J Biol Chem. 2005; 280:13879–13887. PMID:
15691837.
32. Jaisser F, Farman N. Emerging roles of the mineralocorticoid receptor in pathology: toward new paradigms in clinical pharmacology. Pharmacol Rev. 2016; 68:49–75. PMID:
26668301.
33. Breitman M, Zilberberg A, Caspi M, Rosin-Arbesfeld R. The armadillo repeat domain of the APC tumor suppressor protein interacts with Striatin family members. Biochim Biophys Acta. 2008; 1783:1792–1802. PMID:
18502210.
34. Meurs KM, Mauceli E, Lahmers S, Acland GM, White SN, Lindblad-Toh K. Genome-wide association identifies a deletion in the 3' untranslated region of striatin in a canine model of arrhythmogenic right ventricular cardiomyopathy. Hum Genet. 2010; 128:315–324. PMID:
20596727.
35. Fujigaki Y, Sakakima M, Sun Y, Goto T, Ohashi N, Fukasawa H, et al. Immunohistochemical study on caveolin-1alpha in regenerating process of tubular cells in gentamicin-induced acute tubular injury in rats. Virchows Arch. 2007; 450:671–681. PMID:
17464513.
36. Breton S, Lisanti MP, Tyszkowski R, McLaughlin M, Brown D. Basolateral distribution of caveolin-1 in the kidney. Absence from H+-atpase-coated endocytic vesicles in intercalated cells. J Histochem Cytochem. 1998; 46:205–214. PMID:
9446827.
37. Garza AE, Pojoga LH, Moize B, Hafiz WM, Opsasnick LA, Siddiqui WT, et al. Critical role of striatin in blood pressure and vascular responses to dietary sodium intake. Hypertension. 2015; 66:674–680. PMID:
26169051.
38. Gupta T, Connors M, Tan JW, Manosroi W, Ahmed N, Ting PY, et al. Striatin gene polymorphic variants are associated with salt sensitive blood pressure in normotensives and hypertensives. Am J Hypertens. 2017; 31:124–131. PMID:
28985281.
39. Cohen AW, Hnasko R, Schubert W, Lisanti MP. Role of caveolae and caveolins in health and disease. Physiol Rev. 2004; 84:1341–1379. PMID:
15383654.
40. Baudrand R, Gupta N, Garza AE, Vaidya A, Leopold JA, Hopkins PN, et al. Caveolin 1 modulates aldosterone-mediated pathways of glucose and lipid homeostasis. J Am Heart Assoc. 2016; 5:e003845. PMID:
27680666.


 PDF
PDF ePub
ePub Citation
Citation Print
Print



 XML Download
XML Download