

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Brucella is a gram-negative intracellular bacterial pathogen whose infection can cause brucellosis in humans and other animals [12]. Brucella can invade blood and lymphatic vessels through wounds, and bacteria that escape being killed by the host's immune system eventually settle in macrophage cells and propagate, leading to persistent infection [3]. Brucella can also inhibit macrophage apoptosis, thereby evading the host's immune system. The VirB operon of Brucella, which encodes the type IV secretion system (T4SS), is required for its intracellular survival and persistence in the host [4]. However, how Brucella escapes the host's immune defenses, changes the intracellular environment, and invades host cells is incompletely described.
Small ubiquitin-related modifier (SUMO) modification, or SUMOylation, is a type of post-translational protein modification that greatly influences cellular processes [5], including signal transduction, immune recognition, apoptosis, inflammation, and antigen presentation [67]. The SUMO protein family has three isoforms, SUMO1, SUMO2, and SUMO3 that are associated with the following 4 ligase types, E1 activating enzyme, E2 activating enzyme, E2 conjugating enzyme 9 (Ubc9), and E3 ligase, all of which have major roles in protein modification [78]. SUMO1 has an important role in clearing bacteria via the immune system [9].
Brucella infection causes autophagy, inflammation reactions, and apoptosis of the host's macrophages, indicating that the mechanism of Brucella infection must be very complex [10]. However, whether the mechanism involves regulation of macrophages by SUMO or Ubc9 is unknown. Herein, we constructed the following cell lines: 1) a SUMO1 overexpression (O-S) cell line, 2) a Ubc9 overexpression (O-U) cell line, 3) a SUMO1 knockdown (K-S) cell line, and 4) a Ubc9 knockdown (K-U) cell line with which to explore the effects of SUMO1 and Ubc9 expression on inflammation, apoptosis, and the intracellular survival of Brucella melitensis 16M. We also explored the interaction between the T4SS-encoding VirB2 operon of B. melitensis 16M and the SUMO1/Ubc9 proteins in Brucella-infected cells. Our findings provide foundations for future studies on bacterial invasion and immune escape, chronic Brucella infection, and development of vaccines and drugs against persistent Brucella infection.
MATERIALS AND METHODS
Growth conditions for bacterial strains and cells lines
B. melitensis 16M (Chinese Academy of Agricultural Sciences, China) and the B. melitensis 16M△VirB2 mutant strain (Key Laboratory of Zoonosis, Shihezi University, China) were grown on tryptic soy agar (Biosciences, USA). RAW264.7 macrophage cells and HEK-293FT cells were obtained from the Cell Resource Center, IBMS, CAMS/PUMC (China) and were cultured in Dulbecco's Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum (FBS).
Plasmids and protein
The pLEX-green fluorescent protein (GFP) lentiviral overexpression vector, and the pSPAX2 and pMD2.G packaging vectors, were purchased from the Xinjiang Key Laboratory of Animal Epidemic Prevention and Control (China). Lentiviral RNA interference (RNAi) vector, pLL3.7-GFP, and the packaging plasmids RSV-REV and PCMV-VSVG were purchased from Xinjiang Key Laboratory of Animal Epidemic Prevention and Control (China). The pMD19-T cloning vector was purchased from Takara (China). VirB2 protein was obtained from The Key Laboratory of Zoonosis, Shihezi University (China).
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assays
RAW264.7 cells were cultured in 96-well culture plates (104 cells/well). The knockdown (pLL3.7-SUMO-1 and pLL3.7-Ubc9) and overexpressing (pLEX-SUMO-1 and pLEX-Ubc9) plasmids were transfected into RAW264.7 cells. Dimethyl sulfoxide (DMSO; 0.1%) was used as a control. After treatment for 12, 24, 48, or 72 h, MTT (5 mg/mL) was added and the cells incubated for 4 h at 37°C. The supernatants were then removed and 150 μL DMSO was added to each well and the plate incubated at 37°C for 15 min. Absorbance at OD570 was measured for each well.
Construction of SUMO1 and K-U and overexpression vectors
We used the GenBank SUMO1 (NM009460.2) and Ubc9 (NM_001177610) gene sequences to design specific RNAi fragments (Table 1). The polymerase chain reaction (PCR) primers were designed using Premier 5.0 software (Table 2). Real-time PCR (RT-PCR) primers for mouse SUMO1 and Ubc9 genes were designed according to their GenBank sequences (Table 3). Total RNA was isolated from RAW264.7 cells using an RNA extraction kit (CW Biotech, China) and was reverse transcribed into complementary DNA (cDNA) using the HiFi-Script first-chain synthesis kit (CW Biotech). SUMO1 and Ubc9 genes were PCR-amplified using cDNA as the template. Each target gene was individually ligated into pMD19-T and, after transformation into Escherichia coli, positive colonies were picked. The recombinant plasmids containing the SUMO1 or Ubc9 genes were purified. The gene fragments were then isolated by HpaI and XhoI digestion and then individually ligated into the pLEX-MCS overexpression vector to generate pLEX-SUMO-1 and pLEX-Ubc9. We also constructed 2 RNAi vectors, pLL3.7-SUMO-1 and pLL3.7-Ubc9, using a similar subcloning strategy.
Table 1
Small interfering RNA fragments



Generation of stable overexpression and knockdown SUMO1 and Ubc9 cell lines
HEK-293FT cells and RAW264.7 cells were transfected with the lentiviral knockdown or overexpression vectors using Lipofectamine 2000 (Invitrogen, USA) (5 mL of plasmid was added to 2 × 105 cells). The plasmids and cells were mixed thoroughly and placed in a 37°C 5% CO2 incubator. K-S, O-S, SUMO1 overexpression-knockdown (OS-KS), K-U, O-U, and Ubc9 overexpression-knockdown (OU-KU) groups were generated. After incubation for 48 h, total RNA from each group was extracted using an RNA extraction kit (CW Biotech). The cDNA was synthesized from the mRNA by using AMV reverse transcriptase (CW Biotech), following the manufacturer's instructions. The stable cell lines were verified by quantitative RT-PCR (qRT-PCR).
Establishing a cell model for B. melitensis 16M infection
To establish a cell model for B. melitensis 16M or Brucella 16M△VirB infection, phosphate-buffered saline or lipopolysaccharide-pretreated cells were infected with B. melitensis 16M or Brucella 16M△VirB at the logarithmic growth phase with a 100:1 (bacteria: cells) multiplicity of infection. All cell groups (2 × 106 cell/well) were cultured in DMEM containing 10% FBS at 37°C under 5% CO2. After 1 h, gentamicin (30 g/mL, Sigma-Aldrich, USA) was added to the cell medium for 30 min to kill the extracellular Brucella. The cells received different treatments at 0, 4, 8, 12, 24, and 48 h.
Enzyme-linked immunosorbent assay (ELISA) detection of cytokines
At 0, 4, 8, 12, 24, and 48 h after B. melitensis 16M infection, cell culture supernatants from the different treatment groups in 12-well plates were collected by centrifugation at 3,000 × g for 20 min. Cytokine levels (interferon γ [IFNγ] and tumor necrosis factor α [TNFα]) in the supernatants were measured by performing ELISA (R&D, USA) in accordance with the manufacturer's instructions.
Brucella survival in cells subjected to different treatments
At 0, 4, 8, 12, 24, and 48 h post-infection, cells were collected, lysed, and subjected to gradient dilution, which produced concentrations of 101, 102, 103, 104, 105, and 106. The bacteria were then evenly coated with Brucella solid medium and incubated at 37°C for 72 h followed by colony counting.
qRT-PCR
At 0, 4, and 12 h post-infection, total RNA (10 μg) was isolated from each of the different cell groups and reverse transcribed into cDNA following the manufacturer's protocol. β-actin was used as the housekeeping gene. qRT-PCR was performed using a Light-Cycler 480 (Roche Applied Science, Germany) with the SYBR Premix Ex TaqTM kit (Applied Biosystems, USA). The 2−ΔΔCt method was used to calculate the relative expression levels of the genes (Table 3).
Western blotting
At 0, 4, 8, 12, 24, and 48 h post-infection, protein lysate samples were boiled for 10 min in sodium dodecyl sulfate (SDS) buffer. The samples were then separated on a 12% SDS-polyacrylamide gel electrophoresis gel and electro-transferred to a nitrocellulose membrane (Millipore, USA) for 40 min. The membrane was blocked with 5% non-fat milk (Thermo Scientific, USA) for 1 h, and then incubated with rabbit anti-SUMO1 or Ubc9 monoclonal antibodies (Abcam, UK) diluted in Tris-buffered saline with Tween 20 (TBST) buffer for 1 h at 37°C. The membrane was washed with TBST buffer 3 times and incubated with HRP-conjugated mouse anti-rabbit IgG (Bioworld, USA) overnight at 4°C. After 3 washes, the membrane was stained using an HRP-DAB kit (Zhongshan Golden Bridge, China).
RESULTS
SUMO1 and Ubc9 protein levels decrease following B. melitensis 16M infection
The levels of SUMO1 and Ubc9 proteins in cells infected with B. melitensis 16M for 0, 4, 8, 12, 24, and 48 h were determined via western blotting. SUMO1 and Ubc9 levels were decreased from 8 h post-infection onward (Fig. 1), indicating that infection with B. melitensis 16M inhibited the expression of SUMO1 and Ubc9 from this time onward.

Inhibition of cell viability by knockdown and overexpression plasmids in a time-dependent manner
We tested the effects of knockdown and overexpression plasmids on cell viability by performing MTT assays. The knockdown (pLL3.7-SUMO-1 and pLL3.7-Ubc9) and overexpression (pLEX-SUMO-1 and pLEX-Ubc9) plasmids produced a reduction in cell viability from 48 h after transfection onward (Fig. 2).

SUMO1 and Ubc9 gene expression levels in differently treated cells
The relative gene expression levels of SUMO1 and Ubc9 were calculated using the 2−ΔΔCt method (Fig. 3). Compared with the control group, SUMO1 expression in K-S cells was significantly decreased (p < 0.01), with knockdown efficiency reaching 53%. Ubc9 expression in K-U cells was also significantly decreased (p < 0.01), with knockdown efficiency reaching 52%. In the O-S and O-U groups, SUMO1 and Ubc9 expressions were significantly increased after lentiviral infection. The expression levels in the OS-KS and OU-KU groups were also significantly increased (p < 0.05). These results indicate that we successfully constructed an experimental platform that included SUMO1 and Ubc9 knockdown, overexpression, and rescue cell lines.
Fig. 3
Quantitative real-time polymerase chain reaction was performed to analyze the mRNA levels of SUMO1 and Ubc9 in lentivirus-infected RAW264.7 macrophages. Statistical significance is indicated as *p < 0.05 or **p < 0.01.
mRNA, messenger RNA; SUMO1, small ubiquitin-related modifier 1; Ubc9, E2 conjugating enzyme 9; NC, no-treatment cell lines; K-S group, stable SUMO1 knockdown cells; O-S group, cells stably overexpressing SUMO1; OS-KS group, SUMO1 overexpression-knockdown cells; K-U group, stable Ubc9 knockdown cells; O-U group, cells stably overexpressing Ubc9; OU-KU group, Ubc9 overexpression-knockdown cells.

Cytokine production in different RAW264.7 cell treatment groups following infection with B. melitensis 16M
Different treatment groups of RAW264.7 cells produced varying levels of IFNγ and TNFα cytokines at different times after infection with B. melitensis 16M compared with the control group. At 4, 8, 12, 24, and 48 h after Brucella 16M infection, the O-S and O-U groups secreted significantly higher levels of both IFNγ and TNFα compared with the control group (p < 0.05), peaking at 12 h post-infection. In contrast, K-S and K-U cells secreted significantly lower levels of IFNγ (Fig. 4A) and TNFα (Fig. 4B; p < 0.05) at 8, 12, 24, and 48 h post-infection compared with the control group (p < 0.05). Additionally, the OS-KS and OU-KU groups showed similar significant changes at 12 h post-infection (p < 0.05). Collectively, these results indicate that after SUMO1 and Ubc9 silencing, the type 1 T helper (Th1) immune response was decreased in Brucella-infected macrophages. Thus, expressions of SUMO1 and Ubc9 appear to benefit the release of Th1 cytokines.
Fig. 4
Enzyme-linked immunosorbent assay detection of IFNγ (A) and TNFα (B) in the supernatants of different treatment groups of RAW264.7 cells after B. melitensis 16M infection. Brucella-infected-untreated cells acted as the control group. Statistical significance is indicated as *p < 0.05 or **p < 0.01.
IFN, interferon; TNF, tumor necrosis factor; K-S group, stable SUMO1 knockdown cells; O-S group, cells stably overexpressing SUMO1; OS-KS group, SUMO1 overexpression-knockdown cells; K-U group, stable Ubc9 knockdown cells; O-U group, cells stably overexpressing Ubc9; OU-KU group, Ubc9 overexpression-knockdown cells.

Intracellular survival of B. melitensis 16M in RAW264.7 cells after different treatments
We determined whether SUMO1 and Ubc9 affected the intracellular survival of B. melitensis 16M in RAW264.7 cells after different treatments (Fig. 5). Brucella colony forming units (CFU) were counted. From 8 h to 48 h, the O-S and O-U groups had lower B. melitensis 16M log CFU values compared with the control group (no treatment) (p < 0.05), but the K-S and K-U groups had significantly higher values (p < 0.05). The differences between groups were most obvious at 12 h post-infection (p < 0.01). The results for the OS-KS and OU-KU groups were essentially the same as those of the control group. These results indicate that inhibition of SUMO1 and Ubc9 proteins promotes B. melitensis 16M growth and reproduction.
Fig. 5
Intracellular survival of post-infection B. melitensis 16M in different RAW264.7 cell treatment groups. RAW264.7 cells were lysed, and Brucella was coated with tryptic soy agar culture medium and cultured at 37°C for 72 h. B. melitensis 16M numbers were then counted. Brucella-infected untreated cells served as the control group. Statistical significance is indicated as *p < 0.05 or **p < 0.01.
CFU, colony-forming unit; K-S group, stable SUMO1 knockdown cells; O-S group, cells stably overexpressing SUMO1; OS-KS group, SUMO1 overexpression-knockdown cells; K-U group, stable Ubc9 knockdown cells; O-U group, cells stably overexpressing Ubc9; OU-KU group, Ubc9 overexpression-knockdown cells.

Effects of the B. melitensis 16M△VirB2 mutant and B. melitensis 16M LPS on SUMO1 and Ubc9 expression in mouse macrophages
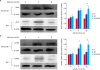
Western blotting and qRT-PCR results showed that incubation of mouse macrophages with Brucella 16M LPS did not induce any change in SUMO1 or Ubc9 expression at any concentration (data not shown). In B. melitensis 16M△VirB2-infected cells, however, western blotting and qRT-PCR indicated that the expression levels of SUMO1 (Fig. 6B) and Ubc9 (Fig. 6A) were significantly increased (p < 0.05) at 4 h and 12 h post-infection compared with the control group. This result indicates that the T4SS-VirB2 can negatively regulate SUMO1 and Ubc9 expressions in macrophages, thereby promoting the intracellular survival of B. melitensis 16M.
Fig. 6
Effect of T4SS-VirB2 on expression of SUMO1 and Ubc9 after infection of RAW264.7 cells with B. melitensis 16M or B. melitensis 16M△VirB mutant. Western blotting was used to detect the protein levels of SUMO1 and Ubc9 (A). Quantitative real-time polymerase chain reaction was conducted to detect the levels of SUMO1 and Ubc9 mRNAs (B). Statistical significance is indicated as *p < 0.05 or **p < 0.01.
NC, no-treatment cell lines; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; mRNA, messenger RNA; SUMO1, small ubiquitin-related modifier 1; Ubc9, E2 conjugating enzyme 9.
VirB2 regulates intracellular survival of Brucella 16M by inhibiting SUMO1 and Ubc9 expression
Western blotting results showed that after incubating cells with VirB protein (from the Brucella 16M IV secretion system, T4SS-VirB), Ubc9 protein levels were decreased at 8 h post-infection (Fig. 7A). This result was similar to that following Brucella 16M infection (Fig. 1). In addition, we observed that intracellular survival of the 16M△VirB2 mutant was significantly decreased at 4 h to 48 h post-infection compared to survival in Brucella 16M infection (p < 0.05; Fig. 7B). A significant increase in colony forming was also observed for VirB2-prestimulated Brucella 16M△VirB2-infected macrophages compared to non-prestimulated Brucella 16M△VirB2-infected macrophages at 12 h and 24 h post-infection. These results indicate that deletion of VirB2 can reduce the virulence of Brucella 16M. The VirB2 protein can regulate the expression of Ubc9, thereby increasing intracellular survival of Brucella in macrophages at the late stage of infection. Thus, T4SS-VirB2 is a key virulence protein that benefits intracellular survival and replication of Brucella 16M.
Fig. 7
VirB2 affects intracellular survival of Brucella 16M by inhibiting Ubc9 expression. Levels of Ubc9 after pre-stimulation with VirB2 for 0, 4, 8, and 24 h were assessed by western blotting (A). Brucella 16M and Brucella 16M△VirB2 cells pre-stimulated or non-prestimulated with VirB2. At 4, 8, 12, 24, and 48 h post infection, the number of bacteria in cells was counted (B). Statistical significance is indicated as *p < 0.05 or **p < 0.01.
CFU, colony-forming unit; Ubc9, E2 conjugating enzyme 9.

DISCUSSION
This study aimed to investigate whether SUMO1 and the enzyme responsible for its conjugation to target proteins, Ubc9, regulate the host immune reaction, apoptosis, and subsequent protection against intracellular survival of Brucella 16M in macrophages. Brucella can maintain a long-lasting interaction with host cells, depending on the secretion of various virulence factors that modulate host cell apoptosis, immune factor production, and antigen presentation [1112]. The Listeria monocytogenes listeriolysin O virulence factor can intentionally or unintentionally interfere with SUMOylation by degrading Ubc9 [13]. SUMOylation can regulate the IFNγ-activated immune response [14]. Considering that Brucella and L. monocytogenes are both intracellular bacteria, their pathogenic mechanisms may be similar; therefore, investigation of the interaction between B. melitensis 16M and SUMO1/Ubc9 in macrophages may help to unravel the invasion and intracellular survival of Brucella 16M. In this study, we confirmed that Brucella 16M can affect the host immune response and host cell apoptosis by interfering with the SUMOylation system (SUMO1/Ubc9). This benefits Brucella intracellular survival at the late stage of infection. VirB2, a key Brucella 16M virulence factor, can decrease SUMO1/Ubc9 expression, thereby helping Brucella 16M avoid death by the host immune system.
SUMOylation has a critical role in many cellular processes, such as immune response and cell proliferation [15]. Cytokines mediate and direct the immune response and are important factors in protection against brucellosis [1617]. In this study, we found that Brucella 16M can strongly activate SUMO1 and Ubc9 expression and can upregulate immune factors at the early stage of infection. Knockdown and overexpression of these two genes directly affected the classical Th1 immune response (IFNγ and TNFα) in Brucella 16M-infected RAW264.7 macrophages (Fig. 4). IFNγ is a key Th1-type immune cytokine that is required for the bactericidal activity of macrophages [18]. TNFα is one of the most important pro-inflammatory and pro-immune cytokines involved in type-1 cell-mediated immune responses to intracellular pathogens, and it positively controls the early production of IL12 and IFNγ in Brucella-infected mice [19]. The absence of TNFα/IFNγ production must favor the early establishment of the bacteria in host cells. Previous studies have suggested that over 90% of Brucella internalized by macrophages are cleared by immune cells at the early stage of infection; however, a few Brucella can survive by evading host immunity and can become established and grow in the endoplasmic reticulum [20]. We suggest that because the host can produce a strong immune response to kill Brucella in the early stage of infection by activating SUMO1/Ubc9, Brucella avoids host immune killing in the late stage of infection by inhibiting SUMO/Ubc9 expression. This reduces the release of SUMO1/Ubc9-mediated immune cytokines and weakens the host's ability to remove Brucella. In this way, Brucella can maintain β-1,2-glucans, the type IV secretion apparatus, and sensing and regulatory systems that allow the bacteria to reach a safe intracellular niche in the endoplasmic reticulum before an effective immune reaction is developed [21]. Given that the humoral immune response, in part mediated bySUMO1/Ubc9, has a pivotal role in controlling Brucella infection, we evaluated the number of intracellular Brucella 16M. The result confirmed that the intracellular survival of Brucella 16M is indeed closely related to the expression of SUMO1/Ubc9. Thus, Brucella improves its intracellular viability by inhibiting the SUMO1/Ubc9-mediated release of immune factors, which also promotes persistence in Brucella infection in vivo.
To further analyze the role of SUMO1/Ubc9 in the survival and proliferation of Brucella 16M in monocytes, we investigated whether Brucella 16M infection affects (induces or prevents) spontaneous or stimulated apoptosis of host cells via the SUMO-mediated release of immune factors. We found that Brucella 16M can reduce the level of IFNγ by inhibiting the expression of SUMO1/Ubc9. Brucella infection renders macrophage-like cells resistant to Fas ligand- or IFNγ-induced apoptosis [22]. This indicates that Brucella not only inhibits the expression of immune factors but also promotes the intracellular survival of bacteria by inhibiting IFNγ-induced apoptosis. Furthermore, previous experiments have confirmed that the release of IFNγ is regulated by SUMO1/Ubc9. By knocking down or overexpressing SUMO1/Ubc9 genes, SUMO1/Ubc9 gradually affected (promoted/inhibited) apoptosis at 24 h after Brucella 16M infection, as shown in Supplementary Fig. 1. Therefore, we suggest that Brucella 16M can reduce apoptosis by reducing SUMO expression at the late stage of infection. Other studies have also reported that SUMO is involved in the regulation of apoptosis [23]. The ability of Brucella to escape host immune killing and survive in cells mainly depends on secreting various effector proteins or virulence factors that inhibit apoptosis and the release of immune factors [11]. Thus, inhibition of apoptosis might represent a beneficial strategy for Brucella development in infected hosts. The result presented in Fig. 4 indicates that the expression of SUMO1/Ubc9 determines the intracellular survival of Brucella 16M, especially in the late stage of infection. Together, our analyses indicate that this is because Brucella infection inhibits SUMO-mediated IFNγ release, thereby inhibiting IFNγ-mediated apoptosis, which promotes the intracellular survival of Brucella. This may be another important cause of chronic Brucella infection in vivo. Previous research attention has focused on revealing the mechanism by which host apoptotic genes are regulated under Brucella infection at proteome and transcriptome levels [24]. However, the mechanism by which Brucella inhibits macrophage apoptosis has not been determined. Therefore, we speculate that our conclusion may help elucidate the relationship between Brucella infection and apoptosis.
However, what components of Brucella 16M affect the expression of SUMO1 and Ubc9? As we know, Brucella 16M is a virulent strain that expresses a complete T4SS and LPS. Brucella LPS, along with 32 other virulence factors, are key to Brucella infection [25]. In this study, Brucella 16M LPS did not induce any changes in SUMO1 or Ubc9 expression in mouse macrophages at any concentration. Thus, LPS did not interact with SUMO1/Ubc9 during Brucella infection. The T4SS complex is comprised of a family of proteins, and with its help, bacteria can secrete effector molecules in host cells when they cross the cell membrane [26]. Brucella, unlike other gram-negative pathogens, does not have a typical type III secretory system. Instead, it secretes virulence factors in host cells mainly through the T4SS complex. The Brucella T4SS complex is comprised of 12 proteins (VirB1–12) that are encoded by VirB operons [26]. VirB2 is a Brucella surface protein. When VirB2 is deleted, the ability of Brucella to survive in mice is significantly reduced [27]. We found that virulence and intracellular viability of B. melitensis 16M△VirB2 were significantly decreased compared those of B. melitensis 16M. Interestingly, expression levels of SUMO1 and Ubc9 were significantly increased in B. melitensis 16M△VirB2 12 h after infection (p < 0.05). We, therefore, speculate that T4SS/VirB2 directly interacts with SUMO and Ubc9, which interferes with SUMOylation. We then incubated macrophages with purified VirB2 protein and observed that VirB2 can activate Ubc9 similarly to that from Brucella 16M in the early stage of infection. A strong immunity can inhibit the development of a persistent Brucella infection [28]. However, in the late stage of infection, VirB2 can inhibit Ubc9 expression. Therefore, we analyzed whether stimulation of VirB2 can rapidly and spontaneously induce a SUMO1/Ubc9-mediated natural immune response to remove the bacteria. We incubated cells with VirB2 and then infected them with the B. melitensis 16M△VirB2 mutant. This improved the intracellular survival of Brucella compared with no VirB2 stimulation. Overall, VirB2 of Brucella T4SS is a key virulence protein that promotes survival and reproduction in host cells by inhibiting SUMO/Ubc9 modification at the late stage of infection.
In conclusions, we report that inhibition of SUMO1/Ubc9 expression by Brucella 16M is a strategy for its intracellular survival. Overexpression of SUMO1 or Ubc9 increases the host's immune response and apoptosis, and it inhibits Brucella 16M survival in macrophages. Silencing of SUMO1 or Ubc9 genes inhibits the host's immune response and apoptosis and promotes Brucella 16M survival in macrophages. We conclude that Brucella 16M can inhibit the release of SUMO-mediated immune factors and apoptosis by reducing the expression of SUMO1 and Ubc9 genes. This reduction promotes escape from immune killing and intracellular survival. In addition, VirB2 may be a key virulence factor of Brucella that inhibits the expression of SUMO1/Ubc9 at the later stage of infection. This study provides a theoretical basis for exploring the molecular mechanism of persistent Brucella infection.
 XML Download
XML Download