

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Xenotransplantation is defined as the process of transplanting organs, tissues, or cells between different species. Porcine organs and tissues have been employed for human organ transplantation in recent years, prompting the use of designated pathogen-free (DPF) pigs as an alternative source for xenotransplantation [1]. However, the presence of bacteria and viruses in porcine tissue continues to pose challenges, with porcine endogenous retrovirus (PERV) being one of the most problematic pathogens [23].
PERV, a member of the Retroviridae family and Gammaretrovirus genus, contains approximately 8 kb of homodimeric RNA genomes similar to other gammaretroviruses [4]. Organization of the PERV viral gene is similar to that of simple retroviruses, consisting of three major viral genes (gag, pol, and env). These structural viral genes are flanked by long terminal repeats (LTRs), which act as viral promoters [5]. The gag gene codes the proteins for matrix, capsid, and nucleocapsid while the pol gene codes reverse transcriptase (RT) and integrase enzymes. The viral envelope glycoprotein is encoded by the env gene, which is made up of surface transmembrane domains [5]. The major sequences of typical retroviral replication can be summarized as infection, formation of proviral DNA, integration to genome, transcription to RNA, packaging, and budding [6]. The repeated sequence blocks harbored in LTRs demonstrate exceptionally strong promotional activity, whereas repeatless LTR can show a reduced effect of viral replication [5].
PERV is integrated at approximately 30 to 50 sites in the genome of different pig breeds [7] and three receptor classes are known, namely PERV-A, -B, and -C [89]. These classes present high sequence homology in the genes for gag and pol, but differ in the genes encoding the envelope proteins (env), which determine the host ranges of the classes [8-10]. The env gene has the motifs or variable regions A and B along with a proline-rich region in the gp70 protein responsible for host tropism [711]. PERV-A and -B were reported to display tropism for the human cell line in vitro [7], while PERV-C-enveloped vectors were shown to have a ‘pig-tropic’ or ecotopic host range [910]. Furthermore, recombinant PERV-A/C demonstrated higher infectivity (500-fold) to human cells than PERV-A in vitro [12]. PERV-A and B can infect human cells in vitro as well as immunosuppressed mice transplanted with pig islets. Fortunately, no PERV transmission has been reported in humans to date; however, no long-term studies in patients under immunosuppression have been reported either [2]. Thus, the risk of infection remains and the deletion or knockdown of PERV sequences could be a solution [13].
Until now, strategies to control replication-competent PERV have involved targeting multiple PERV genes with inhibitory RNA (RNAi) methods [1415]. Although several PERV target genes have been successfully inhibited both at the messenger RNA (mRNA) and protein levels, none have been shown to inhibit PERV infectivity beyond 80%–90% [1617], even when using lentiviral vectors to stably express short hairpin RNAs [1618]. Pigs transgenic for short hairpin RNA (shRNA) targeting the pol gene were generated through somatic cell nuclear transfer, with the live-born piglets showing normal weight and no malformations [18]. The transgene was present in all 6 piglets and the shRNA was detected in all organs. PERV expression was significantly inhibited by up to 94% in all organs of the two transgenic piglets tested [18]. These shRNA vectors can be transfected into primary pig fibroblasts, allowing for the production of PERV-controlled transgenic pigs in which PERV expression can be suppressed for prolonged periods. Additionally, genome-wide inactivation of PERV has been achieved using CRISPR/Cas9 [19], an essential technique for safe xenotransplantation.
In a previous study, the same authors [14] attempted to target the LTR region with a dual LTR 1+2 microRNA (miRNA), which was designed based on the sequences of pigs whose organs were eligible for xenotransplantation. In our previous study [3], we observed a dramatic reduction in PERV expression following LTR inhibition. However, the experimental design for the previous study did not include primary porcine kidney cells and inhibition of the env gene was not investigated [14]. Therefore, this study was performed on primary porcine kidney cells to determine whether miRNAs that target specific regions of the LTR could simultaneously exert an inhibitory effect on the expression of LTR, gag, pol, and env genes.
MATERIALS AND METHODS
miRNA and expression vector design
The primary porcine kidney cells (from the kidneys of 5-day-old piglets) used in this study are the raw materials used for manufacturing vaccines at Green Cross Veterinary Products Co., Ltd. We used the LTR DF and DR primers (Table 1) to identify the LTR sequence for the primary porcine kidney cells. The polymerase chain reaction (PCR) product identified DNA fragments corresponding to 600–650 bp. These DNA samples were used for TA cloning, utilizing a commercial TA cloning kit (TOPcloner™ TA kit; Enzynomics, Korea), and subsequently transformed into competent Escherichia coli cells (DH5a) [14]. Additionally, primary porcine kidney cells were genotyped using the targeting enveloped gene primer sets shown in Table 1 (env AF, env AR, env BF, env BR, env CF, env CR) [20].
Table 1
Primers used for the detection of PERV regions

| Primer | Method | Sequence (5′–3′) | Location* (nt) | Region (Genes) | Size (bp) | Reference |
|---|---|---|---|---|---|---|
| Gapdh F | qPCR | CAGCAATGCCTCCTGTACCA | 776–845 | GAPDH | 70 | [14] |
| Gapdh R | GATGCCGAAGTTGTCATGGA | |||||
| LTR F | qPCR | CCCATAAAAGCTGTCCCAACTC | 523–600 | LTR | 78 | [14] |
| LTR R | GCTGGTGCCCACAGTCGTA | |||||
| Gag F | qPCR | GGTTGCAAAATGAGATTGACATG | 1450–1525 | gag | 76 | [14] |
| Gag R | TCCCTACCTTCAGCCGTGTT | |||||
| Pol F | qPCR | GATCCATGCATCCCACGTTAA | 5615–5684 | pol | 70 | [14] |
| Pol R | ATTTTCAGTCTTTTCGGCTTTCC | |||||
| Env F | qPCR | AAGGCACCTGCATAGGAAAGG | 6671–6741 | env | 71 | This study |
| Env R | TTGATTAAAGGCTTCAGTGTGGTTA | |||||
| HGapdh F | qPCR | GCGCCCCCGGTTTCTATA | 70–148 | GAPDH | 79 | [14] |
| HGapdh R | GATGCGGCTGACTGTCGAA | |||||
| env AF | RT-PCR | TGGAAAGATTGGCAACAGCG | env | 359 | [20] | |
| env AR | AGTGAATGTTAGGCTCAGTGG | |||||
| env BF | RT-PCR | TTCTCCTTTGTCAATTCCGG | env | 263 | [20] | |
| env BR | TACTTTATCGGGTCCCACTG | |||||
| env CF | RT-PCR | CTGACCTGGATTAGAACTGG | env | 281 | [20] | |
| env CR | ATGTTAGAGGATGGTCCTGG | |||||
| LTR DF | PCR | TTCTTGCTGTTTTAGGGCTTG | 1–635 | LTR | 635 | This study |
| LTR DR | AATGAAAGGCCAGTAGAAAGA |
The LTR sequence of the primary porcine kidney cells was used as the template for designing the miRNAs. Based on our previous report [14], dual-targeting (LTR1 + LTR2) miRNAs have better inhibitory effects on the mRNA expression of LTR, gag, and pol genes than single-targeting miRNAs (LTR1 and LTR2). Therefore, we designed a dual LTR 1+2 miRNA based on the methods used in our previously reported study [14], in which we had designed a miRNA vector targeting the 5’LTR U3 region of PERV.
Primary porcine kidney cell culture and transfection
Primary porcine kidney cells were cultured in T-75 flasks with Dulbecco's Modified Eagle Medium supplemented with 10% fetal bovine serum (FBS) and 1% antibiotics for 5 days at 37ºC under 5% CO2. Reverse transfection of miRNAs into primary porcine kidney cells was performed using Lipofectamine 2000 (Invitrogen, USA) according to the manufacturer's instructions. The transfection condition was as follows: primary porcine kidney cells were seeded overnight at a density of 80,000 cells/well (6-well plates). The transfection complex consisted of the miRNA expression vector (1.8 μg) and 12 μL Lipofectamine. Transfected primary porcine kidney cells were maintained in 1X Opti-MEM (Gibco, UK) supplemented with 10% FBS without antibiotics at 37ºC and 5% CO2. The transfection efficiency was automatically measured using the cell transfection program from Bertine Instruments (InCellis®, France).
Generating a stable primary porcine kidney cell clone for constitutive miRNA expression
Blasticidin was used to positively select primary porcine kidney cells containing the miRNA expression vector with a blasticidin-resistant cassette. Briefly, transfected primary porcine kidney cells were seeded in six-well plates and allowed to grow overnight to 60% confluence. The culture medium was replaced the next day with Opti-MEM media containing 12 μg/mL blasticidin (sufficient to kill non-transfected primary porcine kidney cells). Blasticin-resistant cells were sub-cultured every three to four days and blasticidin-resistant colonies were obtained after 20 days and 6 passages. At least 10 blasticidin-resistant colonies (stably transfected; green fluorescent protein continuous expression) per construct were selected to evaluate the knockdown of the target genes.
Measurement of the inhibition of mRNA expression
The knockdown efficiency induced by dual LTR 1+2 miRNA was evaluated based on the expression of LTR, gag, pol, and env regions using real-time quantitative PCR. Total RNA was extracted from transfected primary porcine kidney cells using the RNA Plus Kit (Qiagen Ltd., UK) according to the manufacturer's instructions, followed by measurement of the RNA concentration with a ultraviolet spectrophotometer at 260 nm. The total RNA concentration was adjusted to 500 ng/20 μL. Genomic DNA contamination was removed using DNase I (Invitrogen). Total RNA was converted into cDNA using 10 pmol oligo dTs primer and the MMuLV cDNA synthesis kit (Invitrogen) according to the manufacturer's instructions. Quantitative PCR (qPCR) was performed with Maxima SYBR Green/ROX qPCR Master Mix (Thermo Scientific, USA) in combination with specific primers (Gapdh F and Gapdh R; LTR F and LTR R; Gag F and Gag R; Pol F and Pol R; Env F and Env R) (Table 1). Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as the internal control. All qPCR experiments were performed on the StepOnePlus Real-Time PCR System (Applied Biosystems, USA).
Measurement of inhibition through RT activity
The effect of miRNA on the expression of PERV was also measured based on the RT activity of the primary porcine kidney cells maintained in blasticidin medium (using passages 2, 4, and 6). The C-type RT activity kit (Cavidi, Sweden) was used to quantify the RT activity of the PERV pol gene according to the manufacturer's protocol. Briefly, RT activity was determined for wells with a reading of A405 within the linear range. Additionally, a standard curve for the C-type RT activity kit was obtained through serial dilutions of MMuLV rRT against the concentration of MMuLV present (LOT number 34213).
Inhibition of PERV in primary porcine kidney cells co-cultured with HeLa cell lines
To further investigate the inhibition efficiency of the dual miRNA, we co-cultured the stably expressed miRNA from the primary porcine kidney cells with HeLa cells free of PERV. If the dual miRNA worked properly, the level of PERV gene expression in the HeLa cells would be negligible. Primary porcine kidney cells and HeLa cells (Korean Cell Line Bank No. 10002) were respectively cultured overnight in insert and 24-well carrier plates (Nunc Cell Culture Inserts and Carrier plate, Thermo Scientific) using 10% FBS in Opti-MEM and incubated at 37°C in a 5% CO2 incubator. The primary porcine kidney cells used in this study were transfected with LTR 1+2 miRNA at the sixth passage and showed stable inhibition of PERV. Additionally, primary porcine kidney cells transfected with negative vector were used as the negative control. After one night (a monolayer formed), the insert plate was transferred to a 24-well carrier plate for co-cultivation. The co-cultivation of HeLa and primary porcine kidney cells was maintained for 24, 48, 72, 96, and 120 h. After incubation for the above time periods, the insert plate with primary porcine kidney cells and the supernatant were completely removed from the 24-well carrier plate and the HeLa cells were harvested with trypsin-ethylenediaminetetraacetic acid (0.05%). RNA was extracted from the HeLa cells and RT-PCR was performed to identify the PERV inhibitory effect according to the mRNA level in the co-cultivated human cells. The expression of the target genes was calculated using relative standard curve values between LTR, gag, pol, and env genes and cellular human GAPDH genes using the specific primers shown in Table 1 (HGapdh F and HGapdh R; LTR F and LTR R; Gag F and Gag R; Pol F and Pol R; Env F and Env R). Each gene-expression value was normalized to human GAPDH and presented as copies/μL. All qPCR runs were performed on the StepOnePlus Real-Time PCR System (Applied Biosystems).
RESULTS
Transfection efficiency
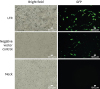
The first part of this study evaluated whether the designed LTR 1+2 miRNA works properly in primary porcine kidney cells. As shown in Fig. 1A, the newly designed miRNA had no mismatches with the LTR sequence of the primary porcine kidney cells. Dual LTR 1+2 miRNAs (targeting nucleotides 130–150 and nucleotides 216–236) for the conserved regions of both primary porcine kidney cell types (GenBank accession numbers FJ357767 and FJ357768) (Fig. 1A) were obtained. With the transfection method used, we were able to achieve 60%–70% transfection efficiency for the LTR 1+2 miRNA vector in primary porcine kidney cells (Fig. 2).
Fig. 1
Confirmation of miRNA targeting site in primary porcine kidney cells and HeLa cells (A) Both LTR1 and LTR2 miRNAs are located at the U3 region of the LTR of PERV [14]. Alignment between miRNAs and the LTR sequences (GenBank accession numbers FJ357767 and FJ357768) was confirmed and primary porcine kidney cells were used for this study. Matches between the LTR1 and LTR2 miRNAs and the LTR sequences of PERV are indicated. (B) Genotyping by RT-PCR in primary porcine kidney cells and HeLa cells using envelop primer sets [20]. No template control.
miRNA, microRNA; LTR, long terminal repeat; PERV, porcine endogenous retrovirus; RT-PCR, reverse transcriptase polymerase chain reaction.

Fig. 2
Determination of miRNA transfection efficiency. Lipofectamine 2000 (Invitrogen) reagent was used to transfect the plasmids (dual LTR 1+2 miRNA, and negative vector control miRNA) into primary porcine kidney cells. Mock-transfected cells were subjected to the transfection process without the addition of miRNA (cells were treated with transfection reagent only). Bright field images (optical microscopy) on the left side and the GFP images (fluorescence microscopy) on the right were taken 24 h post-transfection. The scale bar in the micrographs = 100 µm.
LTR, long terminal repeat; GFP, green fluorescent protein; miRNA, microRNA.
In vitro knockdown of PERV mRNA in primary porcine kidney cells
The primary porcine kidney cells used in this study were confirmed to harbor PERV-B through genotype-specific PCR (Fig. 1B). After the dual LTR 1+2 miRNA was successfully transfected, a decrease in the gene expression level from approximately 20% to 80% (compared to the negative control) was observed in the LTR, gag, pol, and env genes after 24 h (dashed lines, Fig. 3). For the same genes, the level of gene expression inhibition fluctuated over time from 24–120 h post-transfection (Fig. 3). However, in the homologous population of LTR 1+2 miRNA-transfected primary porcine kidney cells (selected with blasticidin), a significant trend of reduced gene expression was observed from passage 2 to passage 6 (Figs. 4 and 5). At the sixth passage, compared to the primary porcine kidney cells transfected with the negative miRNA expression vector, the clone of primary porcine kidney cells that stably expressed dual LTR 1+2 miRNA significantly inhibited the LTR (86.9%), gag (61.2%), and env genes (85.2%) (Fig. 4).
Fig. 3
Knockdown efficiency of miRNA (dual LTR1+LTR2)-transfected primary porcine kidney cells. Real-time PCR was used to detect the expression level of PERV mRNA in primary porcine kidney cells relative to the standard negative vector control (RQ = 100).
miRNA, microRNA; PCR, polymerase chain reaction; PERV, porcine endogenous retrovirus; mRNA, messenger RNA; RQ, relative quantity; LTR, long terminal repeat; SD, standard deviation.

Fig. 4
Selected blasticidin-resistant colonies and analysis of PERV pol gene mRNA levels based on passage numbers. (A) Following transfection with dual LTR 1+2 miRNA, blasticidin-resistant colonies were selected after 20 days at passage 6. (B) Dual LTR 1+2 miRNA targeting the LTR, gag, and env regions. The negative vector-transfected control is regarded as the standard control (RQ = 100 value). Bright field images (optical microscopy) are shown on the left and fluorescence microscopy images (GFP filter) are shown on the right. The scale bar in the micrographs = 100 µm.
PERV, porcine endogenous retrovirus; mRNA, messenger RNA; LTR, long terminal repeat; miRNA, microRNA; GFP, green fluorescent protein; RQ, relative quantity.
*Asterisk marks indicate statistical significance between targeting site groups and negative vector control group.
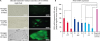
Fig. 5
PERV pol gene analysis by 2 methods: reverse transcriptase (RT) activity and mRNA levels in blasticidin-resistant primary porcine kidney cells over 20 days. (A) Plots show the negative vector control as the standard and sample values are represented by percentage for passages 2, 4, and 6. (B) A standard curve for the C-type RT activity kit was obtained with serial dilutions of MMuLV rRT with respect to the concentration of MMuLV present. The equation for the curve is as follows: y = 1.4551x + 0.0611 (R2 = 0.99). (C) Real-time PCR was used to detect the expression level of the PERV pol gene mRNA in primary porcine kidney cells relative to the standard negative vector control (RQ = 100).
PERV, porcine endogenous retrovirus; RT, reverse transcriptase; mRNA, messenger RNA; PCR, polymerase chain reaction; RQ, relative quantity.
*Asterisk marks indicate statistically significant differences among targeting site groups (p < 0.05) by paired t-test.

RT activity and pol gene expression
We measured the RT activity and mRNA expression of the PERV pol gene. The results showed a clear difference between the RT activity of miRNA- and mock-transfected primary porcine kidney cells (Fig. 5). The RT activity was reduced by 58.7%, 79.2%, and 84.4% in the presence of blasticidin in sub-cultured cells at passages two, four, and six, respectively. Statistical significance was confirmed in passages 2 and 6 with paired t-tests. The mRNA expression level of the pol gene showed patterns that corresponded with RT activity in passage 6, which was the most efficient at 86.9%. Passages 2 and four had RT activity of 53.8% and 76.9%.
Expression of PERV in HeLa cells after co-culture with PERV-inhibited primary porcine kidney cells
HeLa cells (human origin) are known to be free of PERV. To determine the inhibition efficiency of dual miRNA, we measured the gene expression of PERV LTR, gag, pol, and env in HeLa cells after co-culture with stably expressed miRNA from primary porcine kidney cells. PERV expression in the infected HeLa cells was measured with real-time PCR using primers against the PERV LTR, gag, pol, and env regions. Compared to the negative control (Fig. 6, gray lines), the expression of the LTR, gag, pol, and env genes was significantly lower (Fig. 6A-D) in the dual LTR 1+2 miRNA transfected cells, measured by copies/µL. The mRNA copy numbers increased progressively with time. In particular, changing the co-culture incubation time from 48 to 120 h resulted in the largest change in the PERV amount in the negative vector control group (gray lines) contrary to HeLa cells co-cultured with primary porcine cells transfected with dual LTR 1+2 miRNA (orange lines).
Fig. 6
Gene expression of PERV in HeLa cells after co-culture with primary porcine kidney cells stably expressing miRNA. Gray line indicates HeLa cells co-cultured with primary porcine kidney cells transfected with off-target expression vector. Orange line denotes HeLa cells co-cultured with primary porcine kidney cells transfected with dual LTR1 + LTR2 miRNA. Blue line represents naive HeLa cells.
PERV, porcine endogenous retrovirus; miRNA, microRNA; mRNA, messenger RNA.

DISCUSSION
Previous experiments in PK15 cells, which are porcine-derived cells, showed inconsistent results (3–5 nt) at the target sites. However, we observed a dramatic reduction in PERV expression following LTR inhibition. In this study, we wanted to determine whether LTR inhibition could result in the reduction of PERV in primary porcine kidney cells. Therefore, the design for this study was based on primary porcine kidney cells without mismatches for the LTR 1+2 miRNA targeting region (Fig. 1). Genotyping results, as shown in Fig. 1B, indicated that the primary porcine kidney cells have the PERV-B type. We focused on targeting the same LTR region described in a previous report [14].
We confirmed that between the mRNA and RT activity levels, the highest inhibition efficiency was obtained at the sixth passage, similar to the results in our previous study [14]. Over time, untransfected (without blasticidin-resistant gene) LTR 1+2 miRNA in the primary porcine kidney cells were killed, leaving blasticidin-resistant colonies with continuous expression of green fluorescent protein (GFP) and PERV inhibition. In a previous study (using the PK15 cell line), the mRNA expression decreased for LTR (89.7%), gag (82.9%), and pol (92.7%). In addition, the RT activity was inhibited by 91.1%. In this study (using primary porcine kidney cells), the dual LTR 1+2 miRNA simultaneously inhibited the expression of the LTR (86.9%), gag (61.2%), pol (84.4%), and env genes (85.2%). Dual miRNA was able to inhibit the RT activity by up to 86.9%. Therefore, we observed inhibition efficiency up to 86.9%, similarity to the results in our previous study [14] in which PERV genes were inhibited in primary porcine kidney cell colonies after 20 days at the sixth passage. Overall, our results suggested that the LTR 1+2 miRNA targeting the promoter region of PERV (LTR) could knock down several PERV genes in primary porcine kidney cells. In addition, we confirmed the env region of PERV was inhibited as well.
In conclusion, this study confirmed that miRNA targeting the LTR region of PERV could positively inhibit the expression of PERV in primary porcine kidney cells. Dual LTR 1+2 miRNA reduced gene expression for the LTR region as well as the expression of functionally important PERV genes such as gag, pol, and env.
 XML Download
XML Download