

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Oral aspirin challenge is the best method of diagnosing nonsteroidal anti-inflammatory drug (NSAID)-exacerbated respiratory disease (NERD), but this is a time-consuming procedure that results in serious complications in some cases. Therefore, the development of noninvasive biomarkers for easy diagnosis has been attempted to confirm the diagnosis of NERD and prevent unexpected complications of nonsteroidal anti-inflammatory drugs (NSAIDs)/aspirin use in susceptible patients. Using several genetic approaches, including biologically plausible candidate genes and genome-wide association studies (GWAS), more than 100 genetic variants have been identified in association with NERD. Among them, the best mechanistic evidence supports intrinsic dysregulation of the leukotriene (LT)/prostaglandin (PG) pathway, leading to increased recruitment of eosinophils and immune effector cells into the target tissues. These effects are mainly mediated by single nucleotide polymorphisms (SNPs) of the genes that regulate mRNA and protein expression responsible for PG and LT metabolism, i.e., LTC4S,12 ALOX5,34 CYSLTR1,567 CYSLTR2,56 TBX,68 EP2,9 and COX21011 (Table 1).
Table 1
NERD-associated single nucleotide polymorphisms in the genes for cysteinyl LTRs, PG, and thromboxane synthesis and their receptors

NERD, nonsteroidal anti-inflammatory drug (NSAID)-exacerbated respiratory disease; LTR, leukotriene; PG, prostaglandin; AF, attributable fraction; AIA, aspirin-induced asthma; ATA, aspirin-tolerance asthma; MAF, minor allele frequency; OR, odds ratio; CI, confidence interval; LTC4S, leukotriene C4 synthase; ALOX5, arachidonate 5-lipoxygenase; CysLTR1, cysteinyl leukotriene receptor 1; CysLTR2, cysteinyl leukotriene receptor 2; PTGER2, prostaglandin E receptor 2; PTGER3, prostaglandin E receptor 3; PTGER4, prostaglandin E receptor 4; PTGIR, prostaglandin I2 receptor; TBXAS1, thromboxane A synthase 1; TBXA2R, thromboxane A2 receptor; RGS7BP, regulator of G protein signaling 7-binding protein.
![]()
An LTC4S−444A/C promoter SNP (rs730012) is among the most widely reported variants associated with NERD, although its association with NERD across studies is inconsistent.1213141516 Three ALOX5 promoter variants have been shown to be associated with NERD and/or its severity of hyperresponsiveness.41718 However, some results have not been replicated due to small sample sizes, ethnic differences between study populations, or epigenetic changes, for the latter of which there is compelling evidence for a role in NERD.1920
CLINICAL EVIDENCE OF EPIGENETIC COMPONENTS IN NERD
The prevalence of NSAID/aspirin hypersensitivity in adult asthmatics varies depending on the method used for diagnosis. In a recent meta-analysis,21 the prevalence was highest when determined by oral provocation test (adults 21%, children 5%), as compared to verbal history (adults 3%, children 2%). In Korea, 6.2% of 836 adult asthmatics showed a positive responses to oral aspirin challenge tests,22 and 5.8% of 1,173 adult asthmatics showed a positive response for NSAID/aspirin hypersensitivity n histories and/or provocation tests.23 NERD shows less familial aggregation compared with asthma, although the European Network on Aspirin-Induced Asthma found that 6% of NERD patients had a family history of aspirin hypersensitivity.24;a study of 1,344 Turkish patients revealed a 3.7% family history of NSAID/aspirin hypersensitivity,25 suggesting an intermediate genetic background for this condition.
NSAID/aspirin hypersensitivity is more common in women than in men, beginning in adulthood at an average age of 30 years. Once developed, it remains throughout life, although sporadic disappearance of intolerance has been reported.26 However, considerable discordance in clinical manifestations of the disease has been noted in identical twin sisters, suggesting the greater influence of environmental factors.2728 As NERD usually develops in middle age after relatively long-term exposure to NSAIDs, and with a low level of familial aggregation,2429 epigenetic mechanisms may make greater contributions than genetic variations.
GLOBAL CHANGES IN CpG METHYLATION OF NASAL POLYPS FROM SUBJECTS WITH NERD
Epidemiological studies have shown that regular use of NSAIDs reduces the risk of development of at least some cancers.3031 via the well-known targets, including COX-1 and COX-2, and other intracellular pathways, including cell cycle, cell differentiation, apoptosis, and regulation of transcription factors (TFs).32 In our previous in vitro study, DNMT3a and 3b mRNAs were elevated in a mucoepidermoid cell line (NCI-H292) within 1 day after stimulation with medium and high doses of aspirin, while DNMT1 and MeCP2 showed no significant changes in expression (Fig. 1). DNMT3b mRNA was enhanced by stimulation for 2 hours with 0.2 and 2 mM aspirin. DNMT3a mRNA level was increased progressively for up to 3 days by treatment with high-dose aspirin. These data suggest that aspirin may induce global CpG methylation, which may affect gene expression. In contrast, NSAIDs are known induce promoter demethylation of Secreted Protein Acidic and Cysteine Rich (SPARC) by repressing DNMT expression.33 In agreement with the experimental data, an epidemiological study showed that chronic aspirin use may be associated with a lower prevalence of E-cadherin (CDH1) promoter methylation in non-neoplastic gastric mucosa.34
 | Fig. 1Effects of aspirin on DNA methyltransferase genes in the mucoepidermoid NCI-H292 lung cell line. Quantitative real-time polymerase chain reaction assay was conducted in a Smart Cycler instrument, and the relative levels of DNMT3a, DNMT3b, and DNMT1 mRNAs were normalized relative to that of peptidylprolyl isomerase A. The data are representative of 3 consecutive experiments.
*P < 0.05 vs. 0 mM aspirin.
|
There have been few studies on global DNA CpG methylation in NERD. In a genome-wide CpG methylation study of nasal polyps in subjects with NERD and aspirin-tolerant asthma (ATA) patients,35 332 CpG sites on 296 genes were hypomethylated and 158 sites on 141 genes were hypermethylated in NERD (Fig. 2). Thus, the NERD-associated proportion of global differential methylated CpG (DMC) was 1.78% (490/27,587 CpGs analyzed in the test kit), which was about 10 times higher than the proportion of DMC in the bronchial epithelium of patients with atopic asthma (0.19%, 53/27,578 CpGs).36 The 490 DMCs were located on 437 genes and, thus, the global proportion of differentially methylated genes (DMGs) was 3.02% (437/14,457 genes analyzed in the test kit). In silico analysis of the 490 DMCs indicated that 409 CpG sites (83.5%) were on promoter regions with 130 hypermethylated CpGs and 279 hypomethylated CpGs (Fig. 3). In general, hypermethylation of cytosines within CpG islands of promoters causes gene silencing, and hypomethylation triggers active transcription.3738 Thus, differential promoter CpG methylation may affect gene expression levels in nasal polyps of NERD patients, as compared to ATA patients. Ontological classification of the 36,127 genes in AmiGo2 (http://amigo.geneontology.org/amigo/search/ontology) indicated 259 genes in the arachidonate pathways (Table 2). Among them, 66 genes were differentially methylated (25.5%), which was 10 times higher than the global proportion of DMG (3.02%). These observations indicated that the genes in the arachidonate pathways are much more labile to CpG methylation in NERD compared to ATA.
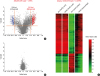 | Fig. 2CpG DNA methylation patterns of nasal polyps and peripheral blood mononuclear cells obtained from subjects with NERD and ATA. Volcano plot of differential methylation levels between NERD and ATA in nasal polyp tissues (A) and buffy coat samples (B). Red dots, delta beta ≥ 0.5 and P ≤ 0.01; blue dots, delta beta ≤ −0.5 and P ≤ 0.01; gray dots, −0.5 ≤ delta beta ≤ 0.5 and P > 0.01. Delta beta, difference in DNA methylation level (subtracting DNA methylation level of ATA from NERD). −log (p), log-transformed t-test P values. (C) Heat map of 490 differentially methylated CpGs between NERD and ATA in buffy coat and nasal polyps. Reproduced with permission from Allergy Asthma Immunol Res 2013;5:258-7619 (license number: EU826007151).NERD, nonsteroidal anti-inflammatory drug (NSAID)-exacerbated respiratory disease; ATA, aspirin-tolerant asthma.
|
 | Fig. 3In silico analysis of the 490 DMCs between NERD and ATA indicated that 409 CpG sites (83.5%) were on promoter regions with 130 hypermethylated CpGs and 279 hypomethylated CpGs.DMC, differential methylated CpG; NERD, nonsteroidal anti-inflammatory drug (NSAID)-exacerbated respiratory disease; ATA, aspirin-tolerant asthma; TSS200, 0–200 bases upstream of the transcription start site; TSS1500, 200–1,500 bases upstream of the transcription start site.
|
Table 2
Proportions of differently methylated and SNP genes in arachidonic acid pathways in NERD compared to those in ATA

Values are presented as number (%). The 259 genes were recruited from 36,127 genes in the AmiGo2, the web-based set of tools for searching and browsing the Gene Ontology database. Number in parenthesis is proportion of genes among the total gene.
NERD, nonsteroidal anti-inflammatory drug (NSAID)-exacerbated respiratory disease; DMG, differentially methylated gene; SNP, single nucleotide polymorphism; ATA, aspirin-tolerance asthma; SNP-gene, number of genes having single nucleotide polymorphism associated with nonsteroidal anti-inflammatory drug (NSAID)-exacerbated respiratory disease; cgSNPs, CpG site related single nucleotide polymorphisms.
*S100A9 (S100 Calcium Binding Protein A9); This protein is a calcium- and zinc-binding protein which plays a prominent role in the regulation of inflammatory processes and immune response.
![]()
GENOME-EPIGENOME INTERACTION VIA CpG SITE-RELATED SNPs (cgSNPs)
Some SNPs influence the presence of CpG sites, where DNA modification, such as methylation and hydroxymethylation, occurs.39 These SNPs can lead to gain or loss of CpG sites and are defined as cgSNPs. For example, a C-to-T transition on the “C” of CpG dinucleotides leads to loss of a CpG site. A human genome study demonstrated a considerable proportion of cgSNPs (23.0%) among 4097556 common variants.40 In the nasal polyps of NERD,35 42 cgSNPs were present in about 11% of the 409 DMC sites on the promoter and one cgSNP was located in the body of the gene (Table 3). Forty-seven TFs were predicted to bind to the DNA sequences of these sites (Fig. 4) on PROMO search (http://alggen.lsi.upc.edu/
). Among the TFs, XBP-1, GR-alpha, and EKNTF-1 bind to more than 10 CpGs sites. The XBP-1 protein is a TF that regulates the expression of genes important to the immune system and the cellular stress response.41 In addition, the expression of this protein is required for transcription of a subset of class II major histocompatibility genes.42 GWAS studies performed by our group and other investigators have revealed that SNPs on HLA-DPB1 (rs1042151 and rs3128965) show the most significant association with NERD susceptibility.43 The rs1042151 acts as a potential cis regulator of the expression of HLA-DPB1 with an expression quantitative trait loci (eQTL) score of 36.83, as calculated using the eQTL browser (http://eqtl.uchicago.edu/cgi-bin/gbrowse/eqtl/). In silico analysis of SNP function indicated that the rs1042151 SNP is located in an exonic splicing enhancer region.44 Thus, XBP-1 is thought to be one of the key TFs binding to the DMCs in the development of NERD, which will be validated in future studies.
Table 3
List of 42 cgSNPs of 409 differentially methylated CpG sites between NERD and ATA and their transcription binding factors

cgSNP, CpG site-related single nucleotide polymorphism; NERD, nonsteroidal anti-inflammatory drug (NSAID)-exacerbated respiratory disease; AIA, aspirin-induced asthma; ATA, aspirin-tolerant asthma; SNP, single nucleotide polymorphism.
![]()
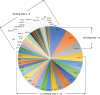 | Fig. 4Diagram of transcription factors binding to 43 CpG site-related single nucleotide polymorphisms.
|
Interestingly, the frequency of cgSNPs was much lower in the arachidonate pathways. Among the total of 259 genes involved in these pathways, the number of genes with SNPs associated with NERD was 49 (18.9%), which was slightly less than the proportion of DMG (25.5%) (Table 3). Seven genes had both DMCs and SNPs. Among them, only one SNP (rs19980990 of S100A9) was related to a CpG site (i.e., a cgSNP). Thus, among the 259 genes, gene expression may be regulated by CpG methylation on 59 genes, by SNPs on 42 genes, and by both CpG methylation and SNPs on 7 genes.
DIFFERENTIALLY METHYLATED CpGs OF THE GENES INVOLVED IN THE PG AND LTR BIOSYNTHESIS PATHWAYS
Among 37 CpGs on 19 genes found in the PG and LT biosynthesis pathways (Supplementary Table S1), there were 14 DMCs on 11 genes, i.e., ALOX12 (cg03760483, cg08946332), ALOX12B (cg03742272), ALOX15 (CG15843823), ALOX15B (CG15799267, CG12343777), ALOX5AP (CG08529529), LOXHD1 (cg17903316), PGDS (cg12554857), PTGDS (CG00563932), PTGES (cg26672426, cg17683775), PTGIS (CG07612655), and TBXAS1 (CG14116569) (Fig. 5). Among them, PGDS (cg12554857) and ALOX5AP (CG08529529) were highly hypomethylated (delta beta: −0.67 and −0.522, respectively), while PTGES (cg26672426) was robustly hypermethylated (delta beta: 0.56) in NERD compared to ATA (Fig. 5). Although the genomic impact of DMCs in NERD has not been analyzed with regard to changes in their genes or metabolites, prostanglandins, cysteinyl LTs, and thromboxanes are expected to be elevated, while PGE may be downregulated by varying the extent of CpG methylation of the relevant genes of NERD patients.
 | Fig. 5Delta beta values of 16 DMCs (P < 0.05) on 11 genes involved in the prostaglandin and leukotriene pathways between NERD and ATA. Delta beta was calculated by subtracting the beta value of ATA from that of NERD at each CpG site.DMC, differential methylated CpG; NERD, nonsteroidal anti-inflammatory drug (NSAID)-exacerbated respiratory disease; ATA, aspirin-tolerant asthma.
|
Therapeutic concentrations of aspirin mainly inhibit cyclooxygenase 1 (COX-1). COX-1, also known as PGG/H synthase 1, PG-endoperoxide synthase 1, or PGH2 synthase 1, is an enzyme encoded in humans by the PTGS1 gene. Therefore, blocking of COX-1 results in a reduction of multifunctional PGH2 level, which is a common precursor substrate for PGD2, PGE2, PGF2, PGI2, and TXB2 (Fig. 6A). Accordingly, the levels of all of these end products are expected to be decreased after aspirin challenge or even in the basal state in NERD. However, systemic PGF245 and PGD2 levels46 and the ratio of local PGD2/PGE2 levels were increased in NERD47 compared to ATA (Fig. 6B-E). These data are in good agreement with the methylation changes of these genes.
 | Fig. 6Effects of aspirin and NSAID on prostaglandin and leukotriene metabolites. Theoretical hanges in PGDs followed by inhibition of COX-1 and actual results (A).47 Changes in PGD2 (B), LTC4 (C), and TXB2 (D) levels in plasma after aspirin challenge in patients with NERD and in those with ATA (license number: 4483431214000).4648NSAID, nonsteroidal anti-inflammatory drug; PGD, prostaglandin D; NERD, nonsteroidal anti-inflammatory drug (NSAID)-exacerbated respiratory disease; ATA, aspirin-tolerant asthma.
*P < 0.05; †P < 0.005.
|
Although systemic basal TBX2 levels were significantly higher in NERD than ATA, the levels were downregulated after aspirin challenge,48 while PGF2 and PGD2 levels were persistently elevated.4546 The different responses to aspirin challenge may be due to genetic differences in these genes between NERD and ATA. A genetic variant study of a Korean population demonstrated that the frequency of the minor allele +141931T>A (rs6962291) in intron 9 of TBXAS1 was significantly lower in the NERD group than the ATA group (Table 1). Taken together, these observations suggest that DNA CpG methylation may exert a regulatory role in synthesis of PG metabolites, especially PGD2 and PGE, while the genetic variants and hypomethylation of TBX2 may be responsible for that of thromboxane B2.49
The other important mechanistic evidence for NERD pathogenesis supports intrinsic dysregulation of the activity of the 5-LO/LTC4S pathway, i.e., LTC4S1 and ALOX54 (Table 1). LTC4S is among the most widely reported variants associated with NERD, although its association with NERD across studies is inconsistent.5051 A recent meta-analysis of 13 case–control studies of asthma revealed significantly increased risk in asthmatic patients carrying the CC or AC genotype vs. the AA genotype.16 Three ALOX5 promoter variants are associated with NERD41718 (Table 1). Interestingly, ALOX5AP showed hypomethylation with delta beta of −0.52, which may affect gene expression. Thus, cysteinyl LTs may be elevated by both differences in the extent of methylation of ALOX5AP and genetic variants of ALOX5 and LTC4 synthase. Among the 14 receptor genes involved in the PG and LTR pathways, 5 genes showed 6 DMC (Supplementary Table S2). However, LTB4R and LXA4 Receptor (FPRL1) are significantly hypomethylated with delta beta > 0.2 (Fig. 7). As shown in Table 1, CysLTR1 and CysLTR2, and PTGER1, PTGER2, PTGER3, and PTGER4, PTGDR, PTGD2R (CRTH2), PTGIR and TBXA2R have different frequencies of SNPs on their respective genes in patients with NERD or those with nasal polyps,52 indicating that the receptors for PG and LTR metabolites have genetic effects due to SNPs except for LTB4R and FPRL1, which are modulated by differences in methylation (Fig. 7).
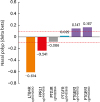 | Fig. 7Delta beta values of 6 DMCs (P < 0.05) on 5 genes of receptors involved in the prostaglandin and leukotriene pathways between NERD and ATA. Delta beta was calculated by subtracting the beta value of ATA from that of NERD at each CpG site.DMC, differential methylated CpG; NERD, nonsteroidal anti-inflammatory drug (NSAID)-exacerbated respiratory disease; ATA, aspirin-tolerant asthma.
|
CONCLUSION
PGE synthesis may be decreased due to hypermethylation of PGEs, while PGD and thromboxane synthesis may be elevated by hypomethylation of PTDS and TBXAS. The imbalance of PGE/PGD may be accentuated by genetic variants of PGERs (EP2–EP4) and PTGDR. In the LT pathways, cysteinyl LTR synthesis may be elevated by hypomethylated ALOX5AP and genetic variants of ALOX5 and LTC4S. Furthermore, the effects of cysteinyl LT may be maximized by genetic variants of CysLR1 and 2 (Fig. 8). Accordingly, the imbalance of PG/cysteinyl LT synthesis may be dominantly regulated by the changes in methylation and may be complemented by the SNPs of LTC4S and ALOX5, and the effects of the imbalance may be accentuated by the genetic variants of PGERs, CYSLTR1, and CYSLTR2 (Fig. 8). Taken together, these observations indicated that subjects with NERD may have genetically distorted and epigenetically susceptible arachidonate pathways to environmental factors, including exposure to NSAIDs or other agents. In addition, DNA methylation may indeed be tightly regulated by genetic factors such as SNPs that affect NERD development (Fig. 8). However, the precise mechanisms by which NSAIDs induce dysregulation of CpG methylation in NERD are still unknown. It remains to be determined when and how the epigenetic effects of NSAIDs begin and the initial mechanisms underlying the changes in CpG methylation. Other possible actions of NSAID may involve modification of other epigenetic components, including histone proteins and miRNA, or altered metabolism of methylating nutrients. Although the molecular mechanisms underlying NERD pathogenesis remain poorly understood, genetic and epigenetic variations play significant roles. Our results enhance the understanding of the genetic and epigenetic mechanisms involved in NERD development and suggest new approaches toward the diagnosis, treatment, and management of NERD.
 | Fig. 8NERD severity and its clinical phenotypes are affected by DNA methylation and genetic variation within genes of associated with multiple pathways for arachidonic acid metabolism.NERD, nonsteroidal anti-inflammatory drug (NSAID)-exacerbated respiratory disease; PG, prostaglandin; SNP, single nucleotide polymorphism; ASA, acetylsalicylic acid ; DP, prostaglandin D (PGD) receptors; EP, prostaglandin E (PGE) receptors; FP, prostaglandin F receptor; IP, prostaglandin I2 receptor; TP, thromboxane receptors; LT, leukotriene; BLT, leukotriene B4 receptor; ALOX5, arachidonate 5-lipoxygenase; 5-HPETE, arachidonic acid 5-hydroperoxide; CysLTR1, cysteinyl leukotriene receptor 1; CysLTR2, cysteinyl leukotriene receptor 2; LTC4S, leukotriene C4 synthase; LTA4H, aeukotriene A4 hydrolase.
|
 XML Download
XML Download