

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Hepatocellular carcinoma (HCC) is one of the most prevalent malignant tumors around the world with the third highest rate of cancer-related mortality, behind only lung cancer and gastric cancer.1 Traditional therapy approaches, such as chemotherapy or radiotherapy, are still the main treatments for HCC. Recently, genomic and immune therapies have emerged as promising, novel treatment options, although they need further investigation.2 Hence, a better understanding of HCC progression at the molecular level will contribute greatly to the development of HCC treatment.
Long non-coding RNAs (lncRNAs) are a group of non-protein-coding RNAs that can regulate gene expression at the chromatin modification, transcriptional, or posttranscriptional level.3 With a length of >200 nucleotides, various lncRNAs have been shown to regulate the development and progression of HCC.4 To date, abnormal expression of many HCC-related lncRNAs has been detected and shown to affect cell metastasis or apoptosis in HCC by targeting corresponding genes.5 Cancer susceptibility candidate 2 (CASC2), located on chromosome 10q26, was first observed to be downregulated in endometrial cancer in 2004 and was identified as a tumor-suppressor in 2007.67 Other studies have reported that CASC2 expression may serve as a clinically utilizable biomarker for outcomes in cancer and melanoma patients.89 In addition, CASC2 exhibits a suppressor role in progression of various tumors, such as osteosarcoma,10 bladder cancer,11 gliomas,12 and HCC.13 Although a large amount of evidence indicates the clinical significance of CASC2 in cancer patient prognosis, its molecular mechanism remains poorly understood.
MicroRNAs (miRNAs), ranging in length from 20 to 22 nucleotides, are small non-coding RNA molecules that are highly conserved in evolution and modulate important cellular processes by directly regulating downstream genes, primarily at the post-transcriptional level.14 It was suggested that certain miRNAs hold the potential to be biomarkers for liver disease and HCC, such as miR-122, miR-125a-5p, miR-1231, miR-18a, miR-221, miR-222, miR-224, miR-101, miR-106b, and miR-195.1516 A growing body of evidence suggests that miRNAs, which function as both oncogenes and tumor suppressors, exert important functions in the development and progression of HCC.17 The hypermethylation of hsa-miR-183 is common in HCC and likely of use as a diagnostic and prognostic marker.18
The Wnt signal transduction cascade regulates various biological processes throughout development, and abnormal Wnt signaling underlies several human diseases.19 The Wnt/β-catenin signaling pathway was reported to participate in the modulation of several malignancies, including colorectal cancers, non-colorectal gastrointestinal cancers, desmoid tumors, breast cancer, adrenocortical tumors, melanoma, glioblastoma multiforme, renal cell carcinoma, osteosarcoma, hematological malignancies, and HCC.2021 In this study, we aimed to evaluate the expression levels of CASC2 and miR-183 in HCC and to confirm the relationship between CASC2 and miR-183, as well as the regulatory mechanism of CASC2 in HCC progression.
MATERIALS AND METHODS
Clinical samples and cell culture
The current study was permitted by the Institutional Review Board of the People's Hospital of Hanchuan, and written informed consent was obtained from 30 HCC patients. A total of 30 pairs of clinical tissues and paired neighboring non-tumor tissues (NTTs) were collected from the 30 patients diagnosed with HCC by means of pathology at the People's Hospital of Hanchuan after surgical resection. None of the patients had received radiotherapy, chemotherapy, or other anticancer therapies. Retrieved specimens were immediately frozen and stored in liquid nitrogen. The patients were traced by telephone or outpatient review to collect their physical condition. Overall survival was defined as the time from surgery to death or the last follow up.
The human normal liver cell line LO2 and five human HCC cell lines, HepG2, Hep3B, QSG-7701, SMMC-7721, and Huh-7, were obtained from American Type Culture Collection (ATCC, Manassas, VA, USA). All cells were maintained in Dulbecco's Modified Eagle Medium (DMEM, Gibco, Grand Island, NY, USA) supplemented with 10% (v/v) fetal bovine serum (FBS, Invitrogen, Carlsbad, CA, USA) in a 5% CO2 humidified incubator at 37℃.
Reagent and cell transfection
The pcDNA3.1 vector was purchased from Thermo Fisher Scientific (Waltham, MA, USA), and pcDNA3.1-CASC2 plasmid was constructed and stored in our laboratory. miR-183 mimic and its negative control (NC) mimic were obtained from GenePharma (Shanghai, China). All of the above oligonucleotides or plasmids were transfected into HCC SMMC-7721 and Huh-7 cells using Lipofectamine™ 2000 reagent (Invitrogen) according to the manufacturer's protocols.
RT-qPCR assay
Total RNA was isolated from cultured cells or HCC tissues using TRIzol reagent (Ambion, Carsland, CA, USA), and 500 ng of total RNA was used as a template for reverse transcription reaction with a ReverTra Ace qPCR RT Kit (Toyobo, Tokyo, Japan). Subsequent qPCR was performed utilizing SYBR Master Mix (Applied Biosystems, Foster City, CA, USA) with StepOne-Plus™ Real-time PCR Systems (Applied Biosystems). β-actin and U6 were used as internal references for CASC2 and miR-183, respectively. The primers for CASC2, miR-183, β-actin, and U6 were as follows: CASC2, 5′-GCACATTGGACGGTGTTTCC-3′ (sense) and 5′-CCCAGTCCTTCACAGGTCAC-3′ (antisense); miR-183, 5′-GTTATTAATAGGAATGGGGTAG-3 (sense) and 5′-AAACRACTCTCAACCTCCC-3′ (anti-sense); β-actin, 5′-ATGGGTCAGAAGGATTCCTATGTG-3′ (sense) and 5′-CTTCATGAGGTAGTCAGTCAGGTC-3′ (anti-sense); U6, 5′-CGCTTCGGCAGCACATATACTA-3 (sense) and 5′-CGCTTCACGAATTTGCGTGTCA-3′ (anti-sense). qPCR experiments for each gene were performed in triplicate. The expression levels of CASC2 and miR-183 were analyzed using the threshold cycle 2−ΔΔCt method.
Colony formation assay
SMMC-7721 and Huh-7 cells after transfection were seeded into 6-well plates (500 cells per well) and then incubated in DMEM with 10% FBS at 37℃ for 2 weeks with replacement of fresh media every 4 days. Colonies were stained with crystal violet (Sigma-Aldrich, St. Louis, MO, USA) for 30 min at room temperature after fixation with 4% paraformaldehyde at 4℃ for 10 min, and the number of colonies with >50 cells were counted. Three independent experiments were conducted.
MTT assay
SMMC-7721 and Huh-7 cells (1×103 per well) were cultured in 96-well plates and incubated for 24 h at 37℃. 20 µL of MTT reagent (5 mg/mL, Sigma-Aldrich) was added into the cell well. After incubation for 4 h at 37℃, the supernatant was removed, and 150 µL of dimethyl sulfoxide (DMSO, Sigma-Aldrich) was added into the cell well. The absorbance of each well was determined at 540 nm by a microplate reader (Bio-Rad, Hercules, CA, USA).
Transwell assay
Cell invasion assays were performed in a 24-well transwell chamber (8-µm pores; BD Biosciences, San Jose, CA, USA) that was coated with Matrigel (BD Biosciences) in advance. SMMC-7721 and Huh-7 cells at a concentration of 2×104 cells/mL were added in the upper chamber (200 µL per well), and the lower chamber was filled with 500 µL of complete medium containing 10% FBS and incubated for 24 h at 37℃. A cotton swab was used to remove the cells on the upper surface of the membrane. Then, the cells on the lower surface were stained with 0.1% crystal violet for 15 min at room temperature, then photographed, and counted. The cell migration assays was performed in similar methods, just without the Matrigel on the membrane.
Western blot assay
Total protein was extracted from HCC SMMC-7721 and Huh-7 cells using a Protein Extraction Kit (Bio-Rad). The concentrations of protein were measured by bicinchoninic acid protein assay kits (BCA, Sigma-Aldrich). Subsequently, 20 µg of proteins were separated by 15% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membranes (Millipore, Billerica, MA, USA). The membranes were blocked in 5% skim milk at room temperature for 2 h. Then, the membranes were incubated with primary antibody against GSP3β (1:1000 dilution), β-catenin (1:1000 dilution), and β-actin (1:2000 dilution) at 4℃ overnight. The next day, the membranes were incubated with matched secondary antibody (1:1000 dilution) at room temperature for 2 h. Finally, protein bands were visualized by electrochemiluminescence system (Pierce Biotechnology, Inc., Rockford, IL, USA). The above antibodies were purchased from Cell Signaling Technology (Danvers, MA, USA).
Luciferase reporter assay
Online software, miRcode (http://www.mircode.org/) and YM500v3 (http://ngs.ym.edu.tw/ym500v2/index.php), were applied to seek the target genes of CASC2, and identified miR-183 as a potential target of CASC2. The wild-type 3′-untranslated region (3′-UTR) of CASC2 containing the putative binding site of miR-183 and its mutant were amplified and subcloned into the psiCHECK-2 luciferase promoter vector (Promega, Madison, WI, USA) to generate wt CASC2 or mutant CASC2 plasmid. Then the SMMC-7721 and Huh-7 cells were co-transfected with wt CASC2 or mutant CASC2 with miR-183 mimic or NC mimic utilizing Lipofectamine™ 2000 reagent (Invitrogen). After 48 h, cells were collected, and a luciferase reporter assay system (Promega) was used to evaluate the luciferase activities according to the manufacturer's instructions. Each experiment was repeated three times.
Statistical analysis
SPSS 18.0 software (SPSS, Chicago, IL, USA) were utilized for statistical analyses. All experiments were performed in triplicate. All data are presented as a mean±standard deviation. Student's t-test was conducted to assess differences between groups with a significance level of p<0.05.
RESULTS
CASC2 significantly downregulated in HCC tissues and cell lines
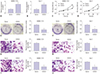
To clarify whether CASC2 participates in the progression of HCC, we first tested the expression level of CASC2 in HCC tumor and matched neighboring NTTs through real-time quantitative PCR (RT-qPCR). The expression levels of CASC2 in 30 HCC specimens were markedly lower than those in paired neighboring NTTs (Fig. 1A). Survival analysis indicated that survival rate in the high CASC2 expression group was much higher than that in the low CASC2 expression group (Fig. 1B). In addition, we measured CASC2 expression in the human normal liver cell line LO2 and five human HCC cell lines (HepG2, Hep3B, QSG-7701, SMMC-7721, and Huh-7). Marked decreases in CASC2 expression were observed in all HCC cell lines (Fig. 1C). Therefore, SMMC-7721 and Huh-7 cells with lower expression of CASC2 were selected for subsequent experiments.
Upregulation of CASC2 inhibits cell viability, colony formation, migration, and invasion in HCC cells in vitro
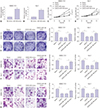
To investigated the role of CASC2 in HCC progression, we constructed SMMC-7721 and Huh-7 cells in which CASC2 was upregulated by transiently transfection with pcDNA3.1-CASC2. Subsequent RT-qPCR assay confirmed the transfection efficiency (Fig. 2A and B). MTT assay showed that cell viability of SMMC-7721 and Huh-7 was decreased after transfection (Fig. 2C and D). Colony formation assay suggested that upregulation of CASC2 significantly inhibited the colony formation ability of SMMC-7721 and Huh-7 (Fig. 2E and F). Subsequent Transwell assay showed that the number of both migratory and invasive cells among SMMC-7721 and Huh-7 sells transfected with pcDNA3.1-CASC2 was less than that in cells transfected with pcDNA3.1 vector, respectively (Fig. 2G–J).
miR-183 is directly targeted by CASC2
The online tools miRcode and YM500v3 were used to predict the downstream targets of CASC2 and showed that the 3′-UTR of CASC2 contains a binding site for miR-183 (Fig. 4A). To further validate the interaction between CASC2 and miR-183, we constructed wild-type (wt CASC2) or mutant CASC2 3′UTR (mut CASC2) with predicted the miR-183 binding site and cloned them into psiCHECK-2 luciferase promoter vector, respectively. These plasmid were co-transfected with miR-183 mimic or NC mimic, after which luciferase activity was tested. Upregulation of miR-183 suppressed the luciferase activity of wt CASC2 in SMMC-7721 and Huh-7 cells, while the luciferase activity of mut CASC2 showed no change (Fig. 4B and C). To verify that CASC2 modulates miR-183, we detected miR- 183 expression in SMMC-7721 and Huh-7 cells overexpressing CASC2 by RT-qPCR. We found that overexpression of CASC2 induced reductions in miR-183 levels and that silencing of CASC2 led to elevations in miR-183 level in the two HCC cell lines (Fig. 4D and E). Pearson analysis indicated that the expression levels of miR-183 were inversely correlated with CASC2 abundance in HCC tissues (Fig. 4F).
Overexpression of miR-183 partially relieves CASC2-mediated inhibition of HCC cell viability, colony formation, migration, and invasion
To further confirm whether CASC2 regulates HCC cells via miR-183, we co-transfected SMMC-7721 and Huh-7 cells with pcDNA3.1-CASC2 or pcDNA3.1 vector with or without miR-183 mimic. After 48 h, RT-qPCR was performed to examine transfection efficiency and showed that miR-183 expression in SMMC-7721 and Huh-7 cells was elevated after transfection with miR-183 mimic (Fig. 5A and B). As exhibited in Fig. 5C and D, upregulation of CASC2 triggered augments in cell viability of SMMC-7721 and Huh-7 cells, while the augment was mitigated by transfection with miR-183 mimic. Similarly, overexpression of CASC2 enhanced the colony formation ability of SMMC-7721 and Huh-7 cells, which was weakened by transfection with miR-183 mimic (Fig. 5E). Transwell assay indicated that miR-183 mimic could, at least in part, rescue the elevation of HCC cell migration or invasion abilities induced by upregulation of CASC2 (Fig. 5F and G).
CASC2 regulates the Wnt/β-catenin pathway by targeting miR-183
To determine the mechanism of CASC2 in the suppression on Wnt/β-catenin signaling pathway, we constructed SMMC-7721 and Huh-7 cells overexpressing CASC2 with or without miR-183 upregulation. Western blot assay was performed to investigate protein levels of C-myc, cyclinD, survivin, and β-catenin in SMMC-7721 and Huh-7 cells. CHIR99021, an agonist of the Wnt/β-catenin pathway, was added in CASC2-overexpressed cells as a positive control. The results indicated that CASC2 upregulation contributes greatly to the activity of Wnt/β-catenin signaling pathway, whereas miR-183 partially reversed the promotion (Fig. 6).
DISCUSSION
The development of HCC typically stems from hepatic cirrhosis caused by various pathogenic factors, including viral hepatitis, non-alcoholic liver diseases, and alcohol abuse.22 More than 600000 people die from HCC per year worldwide.23 It is urgent to investigate the progression of HCC as deep as possible.
Liu, et al.24 analyzed the expression profiles of lncRNAs in HCC using a microarray-based method and found that 659 lncRNAs were differentially expressed between HCC tissues and NTTs. Among these, 171 lncRNAs were significantly downregulated, and 488 were upregulated in HCC tissues, compared to NTTs, indicating that lncRNAs have the potential to serve as therapeutic targets or diagnostic biomarkers.24 Zhu, et al.25 also pointed out that the expression of CASC2 can serve as prognostic factor for cancer patient outcomes. Moreover, Fan, et al.26 validated that CASC2 expression is downregulated in HCC tissues, compared with paired normal tissues, and that the overexpression of lncRNA CASC2 suppresses the viability and triggers apoptosis of HCC cells by targeting miR-24-3p. Also, in HCC, Zhao, et al.14 demonstrated that CASC2 is downregulated in human HCC tissues and HCC cell lines, compared to adjacent non-tumor tissues and a liver cell line, respectively. Moreover, CASC2 overexpression inhibited cell migration and invasion in HCC.14 Here, we found that CASC2 is downregulated in HCC tissues and cell lines and that the introduction of CASC2 inhibits cell viability, colony formation, migration, and invasion in HCC cells in vitro, similar to previous studies. Thus, we sought to identify molecular mechanism (s) by which CASC2 regulates HCC progression.
Dysregulation of miR-183 has been reported in lung cancer, breast cancer, osteosarcoma, and HCC.1827 Here, we evaluated miR-183 expression levels in HCC tissues and cell lines, and found that miR-183 was upregulated in HCC tissues and SMMC-7721 and Huh-7 cells. miR-183 is one member of miR-182–183 miRNA cluster, which comprises miR-96, miR-182, and miR-183.28 Ma, et al.29 highlighted the dual role of this cluster in tumorigenesis, cancer progression, tumor invasion, and metastasis of different tumors. A previous study suggested that miR-183 inhibited proliferation, epithelial-mesenchymal transition (EMT), migration, and invasion of human NSCLC cells by down-regulating MTA1.30 Wang, et al.31 proved that miR-183 targets and downregulates LRP6 expression and the growth, migration, and invasion of retinoblastoma cells. A study conducted by Xu, et al.32 indicated that miR-183 is downregulated in gastric cancer tissues and cell lines and inhibits gastric cancer cell proliferation and invasion by targeting Bmi-1. Also, miR-183 may be a potential therapeutic target for gastric cancer patients.32 In the contrary, in pediatric acute myeloid leukemia, miR-183 acts as a oncogene and facilitates the proliferation and G1/S transition, but inhibits cell apoptosis, in two leukemia cell lines.33 In endometrial cancer, miR-183 was shown to induce EMT, repress apoptosis, and promote cell proliferation, migration, invasion, and in vivo tumorigenesis by targeting CPEB1.34 In our study, we observed marked increases in miR-183 expression in HCC tissues and HCC cell lines. We constructed SMMC-7721 and Huh-7 cells that upregulated CASC2 and examined the resultant cell viability, colony formation, migration, and invasion. Strikingly, overexpression of CASC2 decreased cell viability, colony formation, migration, and invasion of the two cell lines. We also confirmed that miR-183 is a target of CASC2 and negatively regulated by CASC2. Moreover, the inhibition of cell viability and the colony formation, migration, and invasion abilities of HCC cells induced by CASC2 was reversed by miR-183 upregulation, further demonstrating that CASC2 targets miR-183.
Wnt/β-catenin signaling pathway, a highly conserved pathway, regulates important cellular processes, including proliferation, differentiation, migration, genetic stability, apoptosis, and stem cell renewal.20 A study conducted by Pei, et al.11 revealed that the up-regulation of CASC2 reduces bladder cancer cell proliferation and metastasis by decreasing the expressions of β-catenin, C-myc, and cyclin D1 and increasing the expression of E-cadherin. In glioma, CASC2 overexpression was found to exert its repressive effects on gliomas by inhibiting the protein expression of β-catenin, cyclin-D1, and C-Myc in U251 cells.12 In the current study, CASC2 upregulation repressed Wnt/β-catenin signaling through sponging and downregulating miR-183 with the inhibition of C-myc, cyclinD, surviving, and β-catenin expression, which is in line with previous studies, while the gain of miR-183 activated the Wnt/β-catenin signaling pathway.
In summary, we observed the downregulation of CASC2 and upregulation of miR-183 in HCC tissues and cells. Further, upregulation of CASC2 suppressed the cell viability and the colony formation, migration, and invasion abilities of HCC cells in vitro, as well as Wnt/β-catenin signaling. miR-183 was identified as a target of CASC2 and to be negatively regulated by CASC2. Gain of miR-183 was able to elevate HCC cell viability, colony formation, migration, and invasion abilities and activate Wnt/β-catenin signaling pathway, while the overexpression of CASC2 weakened the effects of miR-183. In conclusion, CASC2 repressed cell viability, colony formation, migration, and invasion of HCC cells by sponging miR-183.




 XML Download
XML Download