

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Androgenic alopecia (AGA) is characterized by the gradual miniaturization of hair follicle cells and a shortened growth period (anagen phase) of dermal papilla (DP) cells, which can lead to hair loss in the frontal area of the scalp1. The duration of the anagen phase is a critical determinant of hair length. After repeated hair cycles, the length of new hair becomes shorter and the frequency of hair shaft shedding increases (lag period). These result in hair follicles being depleted of hair shafts and cause the miniaturization of hair follicles, which eventually leads to their loss. These phenomena are mainly mediated by androgens circulating in the blood and the altered androgen sensitivity of hair follicles. DP cells can produce androgen to activate androgen receptor (AR)2. It is known that ARs are only expressed in the DP cells3. The DP cells isolated from balding hair follicles show higher levels of ARs than those isolated from non-balding hair follicles45.
Levels of dihydrotestosterone (DHT) are highly correlated with development of AGA6. DHT is produced from testosterone, and this reaction is catalyzed by cytoplasmic 5α-reductase. Finasteride has been developed to reduce DHT production in the scalp skin by inhibiting 5α-reductase. Compared to testosterone, DHT is a more potent androgen that binds more strongly to the AR. The concentration of DHT is higher in a balding scalp than in a non-balding scalp7. In AGA, DP cells from the androgen-sensitive frontal scalp contain more 5α-reductase type II than those from the non-androgen-sensitive occipital scalp5.
AR signaling is the main pathway involved in the development of AGA in DP cells. After DHT binds to the AR, the AR-DHT complex translocates to the nucleus after dimer formation where AR co-activators are recruited to the AR-DHT complex. This complex induces the expression of AR-driven genes, including transforming growth factor-beta 1 (TGF-β1), Dickkopf-1 (DKK1), and interleukin 6 (IL6), which act as inducers for the premature catagen phase in the hair cycle, leading to apoptosis of outer root sheath keratinocytes and hair growth arrest8910. DKK1 inhibits canonical Wnt/β-catenin signaling to block hair growth and induces the expression of a pro-apoptotic protein Bax, resulting in apoptosis. TGF-β1-induced transcript 1 (TGFβ1I1) can increase androgen sensitivity in balding DP cells. Increased expression of TGF-β1 can also promote apoptosis11. Elevated IL6 levels induce the anagen-to-catagen transition in the hair follicle and result in hair growth inhibition12.
Finasteride has been proven a good therapy for AGA; therefore, 5α-reductase has been admitted as the best target for treatment of AGA. Nevertheless, some reports suggested that other genes such as TGFβ1I1, AR, and neurotrophin-4 (NT-4) could also be candidates for AGA. TGFβ 1I1 acts as a co-activator of AR and was also highly expressed in DP cells of hair follicles in AGA1013. NT-4 is transcriptionally activated by the AR-DHT complex and its inhibitor induced hair regrowth in DHT-implanted mice14. Further, high levels of reactive oxygen species (ROS) were also expected to induce premature senescence of DP cells15. Therefore, further researches to find more effective targets are required to help AGA patients.
This study was conducted to verify the biological activities of the herb extract, RE-ORGA, in preventing hair loss in DHT-treated DP cells. For this purpose, cell viability and cytotoxicity were tested for DP cells, the expression levels of 5α-reductase type II and Bax were examined, and RNA sequencing (RNA-seq) were performed to the mRNA expression profiles of genes influenced by RE-ORGA. Additionally, anti-inflammatory activities of RE-ORGA were assessed through expression levels of tumor necrosis factor-alpha (TNF-α) and IL6 in mast cells.
MATERIALS AND METHODS
RE-ORGA extract preparation
RE-ORGA is an extract obtained from a mixture of various Korean herbs using the steam distillation method. The main herbs in the mixture are Panax ginseng C. A. Meyer (root), Glycine max (seed), Houttuynia cordata (stem and leaf), Lycium chinense (fruit), Glycyrrhiza uralensis (stem), Citrus unshiu Markovich (peel), Zizyphus jujuba Miller (fruit), Perilla frutescens var. acuta (Odash.) Kudo 4 (leaf), Camellia sinensis (leaf), and Cynanchum wilfordii (root). Deionized water (200 ml) was used to obtain the RE-ORGA aqueous extract. After the mixture of Korean herbs (100 g) was soaked and fully immersed in water, the water was boiled for 3 hours. The herb extract was filtered through a 0.45-µm Millipore syringe filter and stored at 4℃.
Cell viability test
Human dermal fibroblasts (ATCC, Manassas, VA, USA) and human DP cells (Promocell, Heidelberg, Germany) were seeded in 96-well plates at a density of 10,000 cells/well and incubated in a humidified atmosphere with air and 5% CO2 at 37℃. These cultured cells were then treated with various concentrations of RE-ORGA for 24 hours. Cell viability was determined by assessing the ability of cells to cleave tetrazolium salt 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyl tetrazolium bromide (MTT) to formazan. Optical densities were measured at a wavelength of 570 nm on a SpectraMax 340PC384 Absorbance Microplate Reader (Molecular Devices, LLC., San Jose, CA, USA).
Cell proliferation test
Cell proliferation was measured using Cell Counting Kit-8 (CCK-8; Dojindo Molecular Technology Inc., Kumamoto, Japan). Human DP cells were seeded in 96-well plates at a density of 3,000 cells/well and incubated in a humidified atmosphere with air and 5% CO2 at 37℃. Cultured cells were then treated with various concentrations of RE-ORGA for 1, 2, and 5 days according to the kit protocol. CCK-8 solution was added, followed by a 4-hour incubation, and absorbance at 450 nm was measured on a SpectraMax 340PC384 Absorbance Microplate Reader.
Western blot analysis
DHT (100 nM) was used to stimulate DP cells to express 5 α-reductase type II and Bax for 5 hours and 48 hours, respectively. Total protein (20 µg) from the human DP cell lysate was separated on NuPAGE Bis-Tris gels (Novex; Life Technologies, San Diego, CA, USA). The proteins were transferred onto a nitrocellulose membrane, followed by incubation with anti-5α-reductase and anti-Bax antibodies (Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA) overnight at 4℃.
RNA sample preparation and Illumina Hiseq2500 sequencing
DP cells were seeded in a 100-mm culture dish (7×105 cells/dish) and grown overnight. They were then treated with 100 nM DHT with or without 20% RE-ORGA for 5 hours. Total RNA was isolated from cells using Trizol (Invitrogen, Carlsbad, CA, USA). The quantity and purity of the extracted RNA were determined by measuring the absorbance at a wavelength of 260 nm with a UV-spectrometer (Nanodrop; Thermo Fisher Scientific, Waltham, MA, USA). cDNA libraries were prepared using the TruSeq RNA Library Preparation Kit (Illumina Inc., San Diego, CA, USA) for 100 bp paired-end sequencing. Briefly, mRNA molecules were purified and fragmented from 2 µg of total RNA using oligo (dT) magnetic beads. The fragmented mRNA was used for synthesis of single-stranded cDNA through random hexamer priming. Using this cDNA as a template for second strand synthesis, double-stranded cDNA was prepared. After a sequential process of end repair, A-tailing, and adapter ligation, the cDNA libraries were amplified using the polymerase chain reaction. The quality of these cDNA libraries was evaluated using an Agilent 2100 BioAnalyzer (Agilent, Santa Clara, CA, USA). The libraries were quantified using the KAPA library quantification kit (Kapa Biosystems, Boston, MA, USA) according to the manufacturer's library quantification protocol. Following cluster amplification of denatured templates, paired-end (2×100 bp) sequencing was performed using the Illumina HiSeq2500 platform (Illumina Inc.).
Inflammatory cytokine measurement
Anti-inflammatory activities of RE-ORGA were evaluated by measuring the levels of TNF-α and IL6 expressed in the human Mast Cell-1 line. Phorbol-12-myristate 13-acetate plus calcium ionophore A23187 (PMACI) was used as a positive control for inflammation. The cells were treated with PMACI and RE-ORGA for 5 hours. Concentrations of TNF-α and IL6 in the cell culture supernatant were determined using commercially available enzyme linked immunosorbent assay (ELISA) kits (Quantikine ELISA Human TNF-α, Cat no.: DTA00C and Quantikine ELISA Human IL6, Cat no.: D6050; R&D Systems, Minneapolis, MN, USA) according to the manufacturer's protocols.
RESULTS
Cytotoxic activities of RE-ORGA
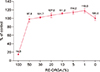
The DP cells were exposed to RE-ORGA at various concentrations (0%~100%), and cell viabilities were tested using the MTT assay. Results are shown in Fig. 1. More than 95% of DP cells survived after treatment with 50% or lower concentration of RE-ORGA. Survival rates of the cells were 100% when treated with RE-ORGA at concentration below 20%. Therefore, the maximum concentration of RE-ORGA was chosen as 20% for further studies.
Cell proliferation assays of RE-ORGA
CCK-8 assays were used to determine whether RE-ORGA could induce proliferation of DP cells. Epidermal growth factor (EGF; 20 ng/ml) was used as a positive control for cell proliferation. As shown in Fig. 2, RE-ORGA at concentrations below 10% promoted growth of DP cells on day 2 and on day 5 compared to the control. However, the number of cells decreased dramatically on all days after treatment with RE-ORGA at concentrations more than 10%. DP cells treated with 20 ng/ml EGF showed significantly higher cell proliferation rate only on day 5 compared to control. When RE-ORGA treated cells were compared with EGF-treated cells, the cells treated with 1% RE-ORGA showed a significantly higher proliferation rate only on day 2 (p-value=0.00005), whereas the cells treated with 5% RE-ORGA showed a significantly higher cell proliferation rate on day 2 (p-value=0.00003) and on day 5 (p-value=0.004). However, cells treated with 10% RE-ORGA showed similar cell proliferation rates as cells treated with EGF at 20 ng/ml.
Western blot analysis on 5α-reductase type II and Bax
Western blotting analysis was performed to examine the expression levels of 5α-reductase type II and Bax in DHT-stimulated DP cells treated with RE-ORGA (Fig. 3). The expression levels of 5α-reductase type II were decreased after treatment with RE-ORGA. After the DHT-stimulated cells were treated with RE-ORGA for 5 hours, the expression levels of 5α-reductase type II decreased to 62.6% in the 5% RE-ORGA treatment group, to 24.1% in the 10% RE-ORGA treatment group, and to 21.3% in the 20% RE-ORGA treatment group compared to their levels in the only DHT-treated cells. Protein levels of Bax (one of the late genes induced by DHT) also decreased after treatment with RE-ORGA. RE-ORGA treatment decreased the expression of 5 α-reductase type II and Bax in a concentration dependent manner. RE-ORGA at 20% resulted in an 86% reduction in the expression of Bax, comparable to the 78% reduction observed after treatment with 200 nM finasteride.
RNA-seq results with RE-ORGA
RNA-seq was performed to investigate changes in expression patterns of AR-induced genes in DHT stimulated DP cells treated with RE-ORGA (Fig. 4). RNA-seq results showed down-regulation of the Bax gene in mRNA levels after treatment with 20% RE-ORGA (log fc: −0.786). However, the mRNA levels for 5α-reductase type II were below the detection limit, even in the cells treated with only DHT. It was found that mRNA transcripts of 5α-reductase type I and 5-α-reductase type III were decreased after RE-ORGA treatment (log fc: −0.158 for 5α-reductase type I, −0.253 for 5-α-reductase type III). RNA-seq revealed that mRNA levels of ARs (log fc: −0.575) and TGF-β1 (log fc: −0.182) were downregulated. In contrast, mRNA levels of DKK1 (log fc: 0.735) and IL6 (log fc: 2.04) were highly increased. It has been reported that two molecules, TGFβ1I1 and protein tyrosine kinase 2 beta (PTK2β), can regulate the transcriptional activity of the AR. RNA seq showed that RE-ORGA induced the expression of PTK2β mRNA (log fc: 0.659) but downregulated the expression of TGFβ1I1 mRNA (log fc: −0.542), indicating that RE-ORGA has a negative effect on the activation of ARs. RE-ORGA also downregulated the mRNA level (log fc: −0.298) of glycogen synthatase kinase 3 beta (GSK3β), which inhibits Wnt signaling via the phosphorylation of β-catenin. However, mRNA level of the β-catenin was almost unaffected by RE-ORGA (log fc: −0.0405).
Anti-inflammation activity of RE-ORGA
We evaluated the anti-inflammatory activities of RE-ORGA by measuring protein levels of TNF-α and IL6 (Fig. 5). Treatment with 40 nM PMACI induced a 4.7-fold (0.28±0.015) increase in the expression levels of TNF-α in human Mast cell-1 when compared with the control (0.06±0.001). When different concentrations (0%~20%) of RE-ORGA were added to cells following PMACI treatment, the levels of TNF-α decreased with increasing concentration of RE-ORGA. The expression levels of TNF-α were reduced to 72.4% (0.20±0.008) in the 20% RE-ORGA treatment group compared to its levels in the 40-nM PMACI-only treatment group. RE-ORGA also decreased the expression of IL6 compared to control, however the decrease in IL6 expression was not dependent on RE-ORGA concentration.
DISCUSSION
RE-ORGA was extracted from a mixture of various Korean herbs by steam distillation. It was prepared to help hair growth in men with AGA. From the results of cytotoxicity tests, 20% of RE-ORGA was chosen as the maximum non-toxic concentration for DP cells. At lower concentrations than 20%, RE-ORGA helped the proliferation of DP cells and its activity was comparable to that of 20 ng/ml EGF. However, RNA-seq was performed with DP cells treated with 20% RE-ORGA to find as many genes that will be influenced by RE-ORGA as possible.
Actually, mRNA levels of 5α-reductase type II were not quantified in RNA-seq analysis. However Western blot analysis showed that RE-ORGA decreased expression of 5α-reductase type II in DHT-stimulated DP cells by RE-ORGA (Fig. 3). Moreover, RNA-seq analyzed that the transcript levels of 5α-reductase type I were reduced by RE-ORGA. A report suggested that AR activation would lead to repress the mRNA expression of 5α-reductase type II but induce the mRNA expression of 5α-reductase type I in prostate cancer cell lines16. Therefore, we assumed that DHT-activated AR had already decreased mRNA levels of 5α-reductase type II in all DP cells.
We had also investigated some candidate genes for AGA and AR-driven genes from RNA-seq results. TGFβ1I1, a gene for co-activator of ARs, was slightly upregulated by DHT but was down-regulated by RE-ORGA. The PTK2β, mRNAs, which act as a negative regulator of ARs, were more activated with RE-ORGA than with DHT. Moreover, only mRNA levels of TGF-β1, among the three main hair-loss effectors (TGF-β1, DKK1, and IL6), was down-regulated with RE-ORGA, while the other two genes were overexpressed. The suppression of TGF-β1 expression with RE-ORGA could help hair loss induced by oxidative stress, because it was reported that DP cells from AGA patients are more sensitive to ROS and that TGF-β1 can be induced by ROS1617. The transcript levels of Bax, a proapoptotic gene, were also down-regulated, which means that RE-ORGA could protect DP cells from apoptosis following shortened anagen phases.
Cytokines may play an important negative role in hair loss by influencing hair growth18. Addition of TNF-α to human hair follicles can dose-dependently inhibit cell growth19. Serum levels of TNF-α in patients with alopecia areata are significantly elevated compared to those in healthy subjects20. IL6 can inhibit hair shaft elongation and suppress proliferation of matrix cells in cultured human hair follicles12. Therefore, we evaluated the anti-inflammatory activity of RE-ORGA in human Mast cell-1 using ELISA assays. The results showed that RE-ORGA could decrease TNF-α expression in dose-dependent manner but not IL6, although these results were not statistically significant. Consequently, it seems that further studies are required to confirm anti-inflammatory activities of RE-ORGA.
Although RE-ORGA can be a new material to help hair loss, we had not yet verified results from RNA-seq analysis in translational levels and we had not revealed the main target proteins of RE-ORGA. Because RE-ORGA is not a single chemical, it is expected to interact with multiple proteins. If all these targets would be revealed through further studies, we could understand the mechanisms for RE-ORGA to lessen hair loss in AGA patients.
In summary, our study showed that RE-ORGA could promote the proliferation of hair follicle DP cells by altering the androgen sensitivity of AR through the transcriptional regulation of TGFβ1I1 gene and PTK2β gene, which might negatively affect the expression of 5α-reductase type I, Bax, TGF-β1, and GSK3β. Anti-inflammatory activities of RE-ORGA might also contribute to the decrease in cytokine-induced cell death. Therefore, we expect that RE-ORGA could be a useful candidate to inhibit hair loss by alleviating progressive AGA induced by male sex hormones.




 XML Download
XML Download