

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Human hair growth cyclically undergoes three distinct stages of active growth (anagen), regression (catagen) and resting (telogen)1. Multiple signaling pathways and genes are involved in regulating the hair growth and cycling2345.
Klotho, named after an ancient Greek goddess of fate, is a putative aging suppressor gene6, increases the life span when it is over-expressed but hastens aging when it is disrupted in mice. The klotho gene with a size of over 50 kb is present on the chromosome 13q12 and contains 5 exons and 4 introns78. Alternative splicing of the klotho RNA gives rise to two portions: one secreted form and another transmembrane form of klotho protein9. The klotho protein plays a crucial role in suppressing aging through modulating various biological signaling pathways. Klotho binds to a cell-surface receptor and inhibits the intracellular signaling of insulin-like growth factor-110. Klotho protein has been reported to bind to multiple Wnt ligands and to repress their ability to activate Wnt signaling11. Doi et al.12 reported that a novel function of secreted klotho is to bind the type I transforming growth factor-beta (TGF-β) receptor and inhibit TGF-β1 binding to cell surface receptors, thereby inhibiting TGF-β1 signaling. Klotho is predominantly existed in the kidney. It is also detectable in the prostate, placenta, pancreas, skeletal muscle, testis, ovary, inner ear and small intestine1013. Defect in klotho gene expression in mice results in phenotypes resembling human premature or accelerated aging syndrome6. Particularly, skin atrophy and hair loss have been observed as consequences of these mutations. These indicate that klotho may act as a modulator in the hair growth.
Klotho expression has only been detected in a few human tissues and cell lines. However, it is unclear whether klotho is expressed in human hair follicles and is associated with hair growth. The aim of this study was to evaluate the expression pattern and role of klotho in human hair follicles.
MATERIALS AND METHODS
Isolation and culture of human hair follicles
This study was approved by the Institutional Ethics Committee of the Yonsei University Wonju College of Medicine (IRB no. YWMR-13-6-053), and all subjects provided were written informed consents. Scalp samples were taken from the occipital areas of healthy subjects who had not prior or current scalp diseases. Hair follicles were separated and cultured as described previously14. Each of the anagen hair follicles was carefully transferred to a 24-well plate (Corning, New York, NY, USA) containing 500 µl of William's E medium (Gibco BRL, Gaithersburg, MD, USA) supplemented with 10 µg/ml insulin (Sigma-Aldrich Inc., St. Louis, MO, USA), 10 ng/ml hydrocortisone (Sigma-Aldrich Inc.), 2 mM L-glutamine (Gibco BRL), 100 IU/ml penicillin and 100 µg/ml streptomycin (Gibco BRL). Test groups were treated with or klotho siRNA (Invitrogen, Carlsbad, CA, USA) or serial dose of klotho protein (R&D Systems, Minneapolis, MN, USA).
Klotho knockdown
Human anagen hair follicles were transfected with klotho siRNA (Invitrogen) or control siRNA (Santa Cruz Biotechnology, Santa Cruz, CA, USA) using transfection reagent (Santa Cruz Biotechnology) at day 0. After transfection, hair follicles were maintained in a 24-well plate with 500 µl William's E medium (Biochrom, Cambridge, UK) supplemented with 10 µg/ml insulin (Sigma-Aldrich Inc.), 10 ng/ml hydrocortisone (Sigma-Aldrich Inc.), 2 mM L-glutamine (Gibco BRL), 100 IU/ml penicillin and 100 µg/ml streptomycin (Gibco BRL). The efficiency of the blockage was evaluated by reverse transcription polymerase chain reaction (RT-PCR) and western blot analysis.
Measurement of hair length and morphology
The length of hair was calculated as the entire length from the hair bulb base to the hair shaft tip using calculating scales attached to the objective lens in the light microscope (Olympus, Tokyo, Japan). Measurements were done at two day intervals until the 8th day of culture and then were statistically evaluated. During measurements, morphology of hair follicle (anagen, early, mid, and late catagen) was seen and the hair cycle score was analyzed as described in previous reports1516. For statistical analysis, hair follicles in anagen VI were referred to a score of 100, early catagen hair follicles as a score of 200, mid catagen hair follicles as a score of 300, and late catagen hair follicles as a score of 400.
RT-PCR analysis
Total RNA was collected and RNA concentration was determined using ultraviolet spectrometer at 260 nm. Aliquots (1.0 µg) of RNA were incubated at 57℃ for 10 minutes with ribonuclease free water. After cooling on ice, primer extension and cDNA synthesis was performed with 1X RT-buffer, 5 mM MgCl2 solution, 1 mM deoxynucleotide triphosphates, 2.5 µM Oligo d(T)16 (Roche, Mannheim, Germany) and 2.5 IU/µl MML-V RTase (Promega, Tokyo, Japan) in a total volume of 20 µl. cDNA proceeded with the following PCR cycling program: 94℃ denaturation for 5 minutes, then 33 cycles of 94℃ for 30 seconds, 59℃ for 30 seconds, 72℃ for 30 seconds, and an extra extension for 5 minutes at 72℃. The following primers were used in this study: human klotho (forward) 5′-ACTCCCCCAGTCAGGTGGCGGTA-3′, (reverse) 5′-TGGGCCCGGGAAACCATTGC TGTC-3′52; human glyceraldehyde 3-phosphate dehydrogenase (forward) 5′-GAAGGTGAAGGTCGGAGT-3′, (reverse) 5′-GAAGATGGTGATGGGATTTC-3′. PCR products were seen on a 2% agarose gel.
Western blot analysis
Total protein was extracted with protein prep kit (Qiagen, Hilden, Germany) and separated by 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred to a polyvinylidene difluoride membrane (Bio-Rad Laboratories, Hercules, CA, USA). The blotted membranes were incubated overnight with primary antibodies against klotho (Abcam, Cambridge, MA, USA) and β-actin (Sigma-Aldrich Inc.) at 4℃. Membranes were then incubated 1 hour with horseradish-peroxidase-conjugated secondary antibodies at room temperature. Protein expression was detected with the ECL system (Santa Cruz Biotechnology).
Immunohistochemical staining
Hair follicles were fixed and embedded in paraffin. Staining was performed on 5 µm paraffin sections. After deparaffinization and rehydration, sections were incubated with a rabbit monoclonal antibody specific for human klotho (1:200; Abcam) for 3 hours at room temperature. Antibody binding was checked by UltraVision LP detection system (Thermo Fisher Scientific Walt, Waltham, MA, USA) according to the manufacturer's instructions. 3-3′ diaminobenzidine (Thermo Fisher Scientific Walt) was used to color development.
Immunofluorescence staining
For Ki-67 staining, sections were incubated with mouse monoclonal Ki-67 antibody (Abcam) and incubated with Alexa Flour 488 labeled donkey anti-mouse secondary antibody (Abcam). Specimens were measured using a Leica TCS-SPE confocal microscope (Leica Microsystems, Bannockburn, IL, USA).
For terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) staining, sections were treated with 20 µg/ml of proteinase K (Millipore Corp., Billerica, MA, USA) for 15 minutes at room temperature and incubated with terminal deoxynucleotidyl transferase enzyme (Millipore corp.) for 60 minutes at room temperature. TUNEL-positive cells were detected by an anti-digoxigenin fluorescein antibody. Specimens were then stained with propidium iodide (Sigma-Aldrich Inc.) and seen using a Leica TCS-SPE confocal microscope (Leica Microsystems). The number of TUNEL positive cells were counted and analyzed.
Statistical analysis
Data management were analyzed by the IBM SPSS Statistics ver. 20.0 (IBM Corp., Armonk, NY, USA) for Windows statistical program. Statistical analysis was done with one-way ANOVA, then by Dunnett's multiple comparison tests to compare three or more groups. The Student's t-test was employed to compare each group. A p-value less than 0.05 was considered to be statistically meaningful. All data are presented as the mean±standard deviation of at least three separate experiments.
RESULTS
Klotho expresses in human hair follicles
The expression of klotho mRNA in hair follicles was identified by RT-PCR analysis with a specific klotho primer pair. A DNA band corresponding to 350 bp was confirmed in human hair follicles (Fig. 1A). The human kidney tissue served as the positive control7, produced a band similar to that in hair follicles (Fig. 1A). Presence of the klotho protein in human hair follicles was identified by western blot analysis with a specific klotho antibody. Klotho protein was detectable in human hair follicles with the expected molecular weight of 116 kDa. Human kidney tissue as the positive control, produced a band similar to that in the hair follicles (Fig. 1B).
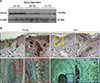
Klotho immunoreactivity is detected in the epithelium of human skin and hair follicles
Immnohistochemical staining was performed on human scalp skin using a polyclonal rabbit antibody against human klotho. Klotho immunoreactivity was detected in both anagen hair follicles (Fig. 2E, E′; F, F′) and extra-follicular structures (Fig. 2A, A′; C, C′; D, D′), while staining was not detected in negative controls (Fig. 2B, B′; G, G′). In human skin, klotho immunoreactivity was mainly expressed in the epithelial cells (Fig. 2A, A′). In human hair follicles, specific immunoreactivity for klotho was found in the outermost areas of outer root sheath (ORS; Fig. 2E, E′). Moderate immunoreactivity for klotho was found in the hair matrix and region of the inner root sheath (Fig. 2F, F′). Klotho positive cells were also found in the sebaceous glands (Fig. 2C, C′) and sweat glands (Fig. 2D, D′). Human kidney tissue, as a positive control, was also expressed klotho in the region of distal convoluted tubules (Fig. 2H, H′).
Klotho expression decreases with aging
To determine the aging dependent expression pattern of klotho in human hair follicles, individual hair follicles (a small piece of scalp skin, aged from 21 to 70) were collected, and klotho expression was identified by western blot assay and immunohistochemistry. Klotho expression was decreased with aging (Fig. 3A) and klotho immunoreactivity was mainly reduced in the epithelial cells of the epidermis and ORS region (Fig. 3B).
Klotho silencing results in hair growth inhibition and premature catagen induction
After transfection with klotho siRNA, hair follicless were cultured over three days. Hair follicle length and hair cycle score were measured. Klotho siRNA treated hair follicles (1,058.58±166.82 µm) had significantly reduced hair growth compared with non-targeted control siRNA treated hair follicles (692.35±99.35 µm) on the third day (Fig. 4A).
Quantitative morphometry of the hair follicles was revealed that klotho silencing accelerates spontaneous catagen developmen (Fig. 4B). It was confirmed by calculation of the hair cycle score (Fig. 4C), which was significantly (p<0.05) increased in the klotho siRNA treated hair follicles (233.74±10.15) compared with the non-targeted control siRNA treated hair follicles (121.27±9.93), indicates that klotho silencing results in premature catagen induction.
Quantitative histomorphometry of TUNEL was revealed that TUNEL positive cells were significantly (p<0.05) increased in klotho siRNA treated hair follicles (43.75±5.93) compared with the controls (23.33±4.16) (Fig.4D, E). However, immunoreactivity of the cell proliferation marker Ki-67 was not significantly changed between klotho siRNA treated hair follicles and the control hair follicles (Fig. 4F, G).
Klotho protein prolongs hair growth and anagen phase
Next, we examined whether treatment with klotho protein resulted in any change in hair growth. Hair follicles were treated with 0.1 µg/ml or 0.5 µg/ml klotho protein every other day and cultured for 8 days. Treatment with klotho significantly (p<0.05) increased hair follicle length (1,764.59±150.59 µm) compared with the control hair follicles (1,427.42±137.78 µm) (Fig. 5A).
DISCUSSION
Klotho has been identified as an “aging-suppressor” gene in mammals that expedites aging upon disruption and boosts that life expectancy upon over-expression917. To date, the expression of klotho has only been partly deciphered. Klotho is expressed in the kidney, brain, prostate, placenta, pancreas, skeletal muscle, testis, ovary, inner ear and small intestine101318. However, it is uncertain whether klotho is presented in human hair follicles. We observed that klotho is expressed in human hair follicles at both gene and protein levels (Fig. 1). In hair follicles, klotho immunoreactivity was mainly observed in the outermost areas of the ORS and hair bulb matrix cells (Fig. 2). Multiple aging-like phenotypes have been first documented in klotho-deficient mice. Skin atrophy and hair loss are also present in this model6. To date, whether klotho expression is correlated with human hair aging has not been clarified. Our results showed that klotho expression is decreased with aging (Fig. 3A) and klotho immunoreactivity is mainly reduced in the epithelial cells of epidermis and ORS region (Fig. 3B). Hair loss and graying are common symptoms of aging1920. Androgenetic alopecia is the most common form of hair loss, which shows clear age-related progression21. Furthermore, some authors have put forward a unique age-related entity, referred to as senescent alopecia2122. Taken together, these results show that klotho may be associated in the regulation of hair aging.
To clarify the functional role of klotho in hair growth and cycling, anagen hair follicles were transfected with human klotho siRNA as described previously1623. In situ knockdown of klotho was observed at the gene and protein levels in organ-cultured hair follicles. Furthermore, we observed that Klotho silencing lead to the premature catagen induction and hair growth inhibition (Fig. 4). We also confirmed that klotho silencing promotes cell apoptosis in follicular keratinocyte (Fig. 4). Otherwise, recombinant klotho protein treatment increases hair follicle length and delays catagen development (Fig. 5). During hair growth phase, multiple factors promoting proliferation, differentiation and survival predominate, whereas hair regression is characterized by activation of various signaling pathways that induce apoptosis in the hair matrix, inner root sheath and ORS keratinocytes2425. Our data suggest that klotho might be involved in the hair cycle through inhibition of cell apoptosis and might operate as a major molecular brake on hair follicle regression.
In conclusion, klotho was expressed in human hair follicles at both gene and protein levels. In hair follicles, klotho expression was prominently observed in the outermost area of ORS and hair bulb matrix cells. Quantification of klotho expression in young and aged donors revealed that klotho immunoreactivity decreased with aging. In hair follicle organ culture, klotho silencing induced premature catagen entering and inhibited hair growth. Otherwise, klotho protein prolonged human hair growth. These results indicate that klotho might be an important regulatory factor for human hair growth and hair cycle change.



 XML Download
XML Download