

 PDF
PDF Citation
Citation Print
Print



Abbreviations
Akt
protein kinase B
AMPK
AMP-activated protein kinase
CIA
collagen-induced arthritis
DC
dendritic cell
IVIg
intravenous Ig
MMP-9
matrix metallopeptidase 9
mTORC1
mTOR complex 1
OC
osteoclast
Pre-OC
osteoclast precursors
RA
rheumatoid arthritis
RANKL
RANK ligand
S6K
ribosomal protein S6 kinase
STAMP
stimulatory transmembrane protein
TRAP
tartrate-resistant acid phosphatase
INTRODUCTION
Many pathological bone diseases are characterized by progressive and excessive bone resorption (1). Therefore, identification of agents to block osteoclast (OC) differentiation and resorption is a common and successful strategy for the development of therapeutic drugs for treatment of OC-related diseases (2).
OCs are specialized bone-resorbing cells regulated by M-CSF and RANK ligand (RANKL) (3).
Th17 cells induce rheumatoid arthritis (RA) on OC activation in published articles by Pöllinger et al. (4) They reported that Th17 cells drive arthritic bone destruction in mice and humans. In our previous studies, Th17 cytokines promoted RANKL production and directly stimulated OC differentiation, causing bone destructive process in RA (56).
Intravenous Ig (IVIg) is a biologic agent with immune modulatory, anti-inflammatory, and immune supplementary effects. It is used as a first-line treatment in patients with Kawasaki disease, dermatomyositis, and immune-mediated thrombocytopenia. IVIg also has extensive off-label indications for various autoimmune diseases (7). In Kawasaki disease, IVIg controls the imbalance of Th17/Treg cells through the downregulation of the number of Th17 cells, and the production of IL-17/Treg cells (8). Th17 induces RA on OC activation. Therefore, there can be potential linkage between IVIg and osteoclastogenesis.
CD4+ T cells, mononuclear phagocytes, fibroblasts, OCs, and neutrophils play major cellular roles in the immunopathology of RA, as does the abnormal production of numerous cytokines, chemokines, and other inflammatory mediators. The autoantibodies contribute directly to the pathogenesis of synovial inflammation and joint destruction via the activation of the complement and Fcγ receptor pathways (9).
Previous research has demonstrated that IVIg suppresses the differentiation and amplification of Th17 cells, as well as the secretion of their effector molecules IL-17, IL-21, IL-22, and CCL20 (1011). These effects of IVIg are associated with inhibition of RAR Related Orphan Receptor C, phosphorylation of STAT3, and reciprocal regulation of Tregs. Since Th17 cells are implicated in the pathogenesis of various diseases where IVIg is also beneficial, our results indicate that inhibition of Th17 cytokine (IL-17, IL-21, and IL-22)–induced osteoclastogenesis represents one of the major mechanisms of action of IVIg. In a previous study, IVIg attenuates tumor necrosis factor-induced bone resorption and it suppressed osteoclastogenesis by induction of A20 expression (12).
In this study, we determined the regulatory effect of IVIg in the Th17 cytokine-induced RANKL expression and OC differentiation, and the inhibitory effect of IVIg in the Th17 differentiation. This study provided important insights into the potential applications of IVIg as a new therapeutic option for preventing bony destructive diseases, such as RA.
MATERIALS AND METHODS
Peripheral blood of healthy donors
Informed consent was obtained from healthy donors, and the experimental protocol was approved by Institutional Review Board (IRB) for Human Research, Konkuk University Medical Center, Seoul, Korea (IRB No. KUH1010186).
Reagents
IL-17, IL-21, IL-22, RANKL, M-CSF, anti-human IL-17, and anti-IFN-γ were obtained from R&D Systems (Minneapolis, MN, USA). IVIg was obtained from IVIg-SN (Green Cross Corp., Yongin, Korea).
Expression of RANKL mRNA by real-time PCR
Peripheral blood monocytes were stimulated with Th17 cytokines. Monocytes were stimulated with or without IVIg for 3 h before treatment of Th17 cytokines. After 72 h, mRNA was measured using real-time PCR as before (13).
Cell isolation from blood
PBMCs were collected from healthy blood by density gradient separation. The monocytes (OC precursors) were prepared from PBMC.
Flow cytometric protocol
Cells were stained to detect with mAbs to CD4, CD25, IL-17, IFN-γ, and Foxp3 (eBioscience, San Diego, CA, USA). Cells were analyzed on a FlowJo software (Tree Star, Ashland, OR, USA).
ELISA
IL-17 and IFN-γ levels in the culture supernatants from PBMCs were measured using sandwich ELISA according to the manufacturer's instructions.
OC formation
Monocytes were pretreated with IVIg for 1 h, after which monocytes were added to each well along with Th17 cytokines. RANKL was used as the positive control. For coculture experiments with CD4+ T cells, human monocytes were seeded in 48-well plates at 5×104 cells/well with 1 ml of medium, and pre-incubated for 1 h with IVIg (0.1, 1, 10, and 20 mg/ml). On day 21, tartrate-resistant acid phosphatase (TRAP)-positive cells were identified as before (13).
Western blot analysis in CD4+ T cells
CD4+ T cells were isolated from the PBMCs as described earlier (16). For the effect of IVIg, CD4+ T cells were pretreated with or without IVIg (10 mg/ml) and cocultured with Th17-differentiation conditions for 1 h. The first Abs to phosphorylate mTOR, AMP-activated protein kinase (AMPK), protein kinase B (Akt), ribosomal protein S6 kinase (S6K), STAT3 (727), and STAT3 (705) (Cell Signaling Technology Inc., Danvers, MA, USA) were diluted in buffers (13).
RESULTS
IVIg reduced Th17 cytokine–induced RANKL expression in CD14+ monocytes
To investigate the suppressive effect of IVIg in Th17 cytokine-induced RANKL expression, CD14+ monocytes were incubated with IVIg (10 mg/ml) for 1 h. The CD14+ monocytes were then cultured with Th17 cytokines, such as IL-17, IL-21, and IL-22 for 72 h. The Th17 cytokines increased the expression of RANKL mRNA. IVIg reduced the Th17 cytokines–induced expression of RANKL mRNA (Fig. 1).

The inhibitory effect of IVIg in Th17 cytokine–induced OC differentiation
The isolated CD14+ monocytes were cultured with RANKL and M-CSF. After 21 days of culture, OCs were differentiated from their precursors. However, pre-incubation with IVIg significantly reduced the RANKL-induced osteoclastogenesis (Fig. 2A). To evaluate the effects of IVIg on Th17 cytokine–induced OC differentiation, isolated CD14+ monocytes were cultured with IL-17, IL-21, IL-22, and M-CSF in the absence of RANKL. After 21 days of culture, OCs were differentiated in the Th17 cytokines and M-CSF culture system. However, IVIg significantly decreased the Th17 cytokine-induced osteoclastogenesis (Fig. 2B-D).
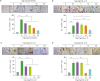
The regulatory effect of IVIg in Th17 cell–induced OC differentiation
When CD14+ monocytes were cultured with Th17 cells in the presence of M-CSF, OCs were differentiated and IVIg decreased the OC differentiation induced by Th17 cells in a dose-dependent manner (Fig. 3A). The gene expression of OC markers, such as TRAP, calcitonin receptor, matrix metallopeptidase 9, cathepsin K, NFATc1, OC-stimulatory transmembrane protein (STAMP) and dendritic cell (DC)-STAMP was increased with Th17 cell addition but their gene expression was reduced by IVIg treatment (Fig. 3B).
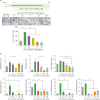 | Figure 3The effect of IVIg in OC differentiation by Th17 condition prestimulated CD4+ T cells. CD14+ monocytes were pretreated. (A) CD4+ T cells were pretreated with IVIg for 1 h, then cultured with Th17 condition for 72 h and then cocultured with CD14+ monocytes in the presence of M-CSF. (B) The gene expression of TRAP, calcitonin receptor, MMP-9, NFATc1, cathepsin K, OC-STAMP, and DC-STAMP from differentiated OCs was measured by real-time PCR. Data were normalized to beta-actin and reported in relative expression units.DC, dendritic cell.
*p<0.05; **p<0.01; ***p<0.001.
|
Suppressive effect of IVIg on Th17 cell polarizing
The proportions of Th17 (CD4+ IL-17+), Th1 (CD4+ IFN-γ+), and Treg (CD25high Foxp3+) cells out of the total CD4+ T cell population are presented in Fig. 4A. Both Th17 cells and Th1 cells were differentiated in Th17 condition. IVIg inhibited the differentiation of IL-17 and IFN-γ expressing cells; however, IVIg did not affect the differentiation of Foxp3 expressing Treg cells (Fig. 4B). IVIg also reduced the production of IL-17 and IFN-γ in the culture medium (Fig. 4C).
 | Figure 4Effect of IVIg on CD4+ T cells isolated from the PBMCs of healthy donors and cultured under Th17-polarizing conditions. Human PBMCs (n=6) were isolated from healthy subjects and pre-incubated with IVIg (0.1, 1, 10, and 20 mg/ml) for 1 h, and then cultured under Th17-polarizing conditions for 72 h. (A) CD4+ T cells were gated for further analysis. Next, the percentage of (B) IL-17+/CD4+ T cells, IFN-γ+/CD4+ T cells, and CD25highfoxp3+/CD4+ T cells was measured by flow cytometry. The production of (C) IL-17 and IFN-γ by Th17-polarizing CD4+ T cells and secretion into the culture supernatant by ELISA.
*p<0.05; **p<0.01; ***p<0.001.
|
The signaling molecules involved in the regulation of IVIg in Th17 polarization
To investigate the signaling involved in the regulation of IVIg in Th17 cell differentiation, CD4+ T cells were incubated with Th17 condition and IVIg. As shown in Fig. 5, the phosphorylation of mTOR, Akt, and S6K increased significantly under Th17 conditions. IVIg significantly inhibited the level of phosphorylated mTOR, its downstream protein pS6K, and Akt, compared with Th17 condition (p-value <0.05 for each).
 | Figure 5Effects of IVIg on the expression of mTOR and AMPK proteins on CD4+ T cells isolated from the PBMCs of healthy donors and cultured under Th17-polarizing conditions. (A) Immunoblotting of p-mTOR, mTOR, p-AMPK, AMPK, p-Akt, Akt, p-S6K, p-STAT3(727), p-STAT3(705), and STAT3 in the CD4+ T cells pretreated with IVIg (10 mg/ml) and then cultured under Th17-differentiation conditions for 1 h. (B) Stimulation of CD4+ T cells under Th17-differentiation conditions, activated phosphorylation of mTOR, Akt, S6K, STAT3 (705), and STAT3 (727) as detected by Western blotting and shown by the ratio of phosphorylated proteins to total proteins.
*p<0.05; **p<0.01; ***p<0.001.
|
DISCUSSION
In this study, we found that IVIg reduced Th17 cytokine–induced RANKL expression and protein production in CD14+ monocytes. In a previous study that we recently conducted, we found that other Th17 cytokines, such as IL-21 and IL-22, induce RANKL expression in RA synovial fibroblasts (6). RANKL and M-CSF are important cytokines in OC formation from the precursor cells (17). This result suggests that IVIg indirectly interrupts the initial process of the Th17 cytokine-associated bony destructive pathways through the inhibition of RANKL production.
In addition to the indirect effect of IVIg on osteoclastogenesis, we determined the direct effect of IVIg in OC formation from their precursors. IVIg directly inhibited the RANKL and Th17-cytokine induced OC differentiation in a dose-dependent manner. This result suggests that IVIg can directly ameliorate the cytokine-induced osteoclastogenesis and that there is a possibility for using it therapeutically.
Next, we determined whether IVIg modulates Th17 cells in direct stimulation of OC differentiation. When monocytes were cocultured with Th17 condition, OC differentiation was increased compared to when they were cocultured with CD4+ T cells. Without RANKL, Th17 cells directly induced OC differentiation and IVIg inhibited the Th17 cell induced osteoclastogenesis. The final goal of treatment in patients with RA is the prevention of joint destruction. Bone erosion caused by activated OCs induces joint destruction and patients’ disability. These results suggest that IVIg can prevent bony destructive processes in RA.
To determine the relationship between IVIg and Th17, we investigated the regulatory role of IVIg in Th17 cell differentiation. IVIg downregulated the number of Th17 cells and IL-17 production, without affecting the number of Treg cells. IVIg reduced osteoclastogenesis via 3 mechanisms. First, IVIg reduced Th17 cytokine–induced RANKL expression in monocytes. Second, IVIg directly inhibited Th17 cytokine–induced OC differentiation from peripheral blood-derived monocytes. Third, IVIg downregulated Th17 cell differentiation. In RA, the clinical efficacy and safety of anti-IL-17 monoclonal Abs, such as secukinumab and ixekizumab, have been under clinical investigation (18). Although dosage does not show the effectiveness of IVIg in clinical trials of low-dose IVIg among patients with RA, the combination therapy of IVIg and IL-17 inhibitors could be a new therapeutic option in patients with severe RA (19).
The previous study and our study share similar results which suggest “IVIg controls the imbalance of Th17/Treg polarization.” However, we did not find IVIg promotes Treg polarization. There are several differences between the 2 studies: 1) We used human peripheral CD4+ T cells, however, they used mouse splenocytes; 2) We used T cells of healthy donors, however they used splenocytes of diseased mouse “collagen-induced arthritis (CIA)”; and 3) Splenocytes from CIA is already exposed by immunologic stimulation, so the focus of immune regulation is both decreasing Th17 response and increasing regulatory system. However, in healthy condition, the focus of immune regulation could be only prevention of abnormal immune response such as Th17 polarization. It does not need to perform regulatory process (20).
Next, we investigated the molecular signaling pathway involved in the suppressive effects of IVIg with CD4+ T cell. We focused on the mTOR/Akt/S6K pathway, which has been shown to have an essential role in the development and differentiation of Th17 cells in many previous studies, as well as in a previous report that we published (2021). We also investigated the activity of Akt and p-S6K, which are located either upstream or downstream of the mTOR/STAT3 pathway, and are significantly associated with mTOR activity (2223). This result showed that the enhanced suppressive effect of IVIg was mediated by an effective regulation of mTOR/Akt/S6K signaling. PI3K/mTOR signaling has been shown to promote effector T-cell activation and differentiation, and our group and others have demonstrated the reorganization of this pathway during the differentiation of naive T-cells into functional suppressor Tregs (2425). Indeed, T-cell specific loss of mTOR correlates with a diminished generation of effector Th1, Th2, and Th17 cells, and enhanced generation of Tregs. Tregs play a major role in the prevention of autoimmunity by suppressing T-cell responses to self-Ags and by limiting the response to foreign Ags.
Additionally, IVIg activates AMPK, which functions to counteract the mTOR signal. In RA, inhibition of mTOR complex 1 (mTORC1) alters the ability of IL-17 to induce mTORC1-dependent proliferation of RA synovial fibroblasts (26).
In conclusion, we found that IVIg significantly suppressed Th17 cytokine–induced RANKL mRNA level and osteoclastogenesis and attenuated Th17 differentiation. Our findings provide important insights into the potential applications of IVIg as a new therapeutic option for preventing OC-associated bony destructive diseases, such as RA.
 XML Download
XML Download