

 PDF
PDF Citation
Citation Print
Print



Abbreviations
Acr
α-crystallin
Ags
antigens
AM
arabinomannan
BCG
Bacillus Calmette Guerin
CFU
colony-forming unit
ESAT-6
early secreted antigenic target of 6 kDa
HBHA
heparin-binding haemagglutinin
LAM
lipoarabinomannan
LTBI
latent tuberculosis infection
mAb
monoclonal Ab
MTB
Mycobacterium tuberculosis
TB
tuberculosis
INTRODUCTION
Tuberculosis (TB) overtook HIV as the biggest killer among infectious diseases caused by a single pathogen, killing 1.3 million people in 2017 (1). The Bacillus Calmette Guerin (BCG) vaccine is largely ineffective and efforts to develop a better vaccine are gathering pace (2). However, a major obstacle in these efforts is the lack of understanding of what makes protective immunity in TB and consequently, the lack of correlates of protection in vaccine design. Cellular immunity is known to contribute significantly to the control of Mycobacterium tuberculosis (MTB) infection (3) but the role of humoral immunity remains contentious. In fact, until a few years ago, the overwhelmingly prevailing view was that Abs have no role to play in immunity to TB.
MTB is considered to be a facultative intracellular pathogen residing predominantly in lung alveolar macrophages. It is for that reason that it was thought that Abs could not gain access to MTB bacilli and therefore could not play a significant role in protection. But what makes an intracellular pathogen? Is any pathogen ever truly intracellular? For example, nobody ever questions the role of Abs against viruses, the obligatory intracellular pathogens. Viruses can only multiply inside a living cell and can only survive but not multiply extracellularly for a limited period of time. Yet, it is in this extracellular phase that they are most vulnerable to Abs. Indeed, most of the protective anti-viral immunity is based on neutralising or blocking Abs, either during the initial infection or subsequent viremia. So, what is the difference to other, non-viral intracellular pathogens, such as bacteria or parasites? Although they reside and multiply within cells, they too, just like viruses, have to leave the infected cell at some point and look for a new host cell. They too could be vulnerable to the interception by Abs and although the outcomes of those interactions are yet to be fully understood, this serves to illustrate that no pathogen is ever exclusively intracellular. It also serves to illustrate that the current dogma that Abs cannot play a protective role against the so-called intracellular bacterial pathogens, whether they be obligatory or facultative, is not evidence based but born of the lack of sufficient understanding of the pathogenesis of the infection. In this article, it is our intention to review the current state of knowledge about the role of Abs in TB, focusing in particular on the progress that has been made in the last 5 years. The reader is further directed to other recent reviews (45) listing various Abs that have been used against MTB and the proposed mechanisms of action.
THE HISTORIC PERSPECTIVE
Ab based immunotherapy was initially termed “serum therapy” and was first developed by Behring and Kitasato in the 1880s. Sera were derived from either vaccinated humans or animals, and the polyclonal antisera were used to treat a range of diseases. The term polyclonal is used as the antisera contained a mixture of different Abs which bind to a number of epitopes of an Ag, or different Ags altogether. Due to issues with purity and the polyclonal nature of the Abs, serum therapy in its infancy had variable outcomes and a wide range of side effects. These side effects were minimised with the subsequent use of monoclonal Abs (mAbs) produced by hybridoma technology and most recently, fully humanised Abs that can be produced at a commercially viable scale. Nowadays, mAb therapies are economically very significant, accounting for some of the highest selling biotechnology drugs (6). However, most of these mAbs are used for non-communicable diseases and relatively much less progress has been made with mAbs for infectious diseases. This can be largely attributed to the rise of sulphonamide antibiotics before the development of hybridoma technology and the obvious cost/effectiveness implications. However, the increasing threat of drug resistance today has renewed interest in the use of mAbs to treat drug resistant infections, for which the treatment options are arduous and limited. An example of such resistant infection is the multi-drug resistant TB, the incidence of which is increasing globally at an alarming rate.
Outside the immunotherapy context, understanding of the role of Abs in protection against MTB is even more important, for TB is a disease in a desperate need of a better vaccine than BCG if the current global epidemic is to be arrested. The BCG vaccine, which shows variably efficacy around the world ranging from 0%–80% (7), is thought to rely solely on induction of cellular immunity against mycobacteria for protection. Indeed, alongside the zoster vaccine (8), it is the only vaccine to do so, among hundreds of vaccines currently in use around the world. Both BCG and its pathogenic relative MTB induce humoral immune responses but they tend to be muted and unreliable as a potential diagnostic target, in contrast to many other infectious diseases. This leads one to speculate that mycobacteria may actively manipulate the host's immune response and skew it towards bacterial growth restricting but ultimately ineffective cell-mediated pro-inflammatory phenotype, as a ploy to avert the potentially damaging action of Abs. Be that as it may, it is now clear that any future TB vaccine design will do well to consider the potential value of inducing a functional Ab response alongside the strong cell mediated immunity. That is why the currently escalating interest in the role of Abs in TB is critical to the conclusion of this long-standing debate in the scientific community and their recognition as a protective element in anti-TB immunity.
EMERGING AND RE-EMERGING THEMES
Several important contributions have been made in recent years towards better understanding of the role of Abs in MTB infection. While the experimental approaches and models of infection differed, several important themes have emerged from these studies, as summarised in Table 1. While some of these themes are entirely new (themes 1 and 2), others draw their origins from previous historical evidence but are re-emerging now with a new impetus for their further experimental exploration utilising novel tools, concepts and models of infection. Coupled with further detailed investigation of the mechanisms of protection involved, these themes offer a unique platform on which to build on our current knowledge and continue advancing this field of research.
Table 1
Emerging themes for the role of Abs in TB

| No. | Emerging theme | References |
|---|---|---|
| 1 | Abs from latently infected individuals are more protective than those from TB patients | (91113) |
| 2 | Ab glycosylation profiles may be associated with different cellular functions | (915) |
| 3 | Ab isotype may play a role possibly dependent on stage of infection and microenvironment | (13161825262730) |
| 4 | Abs are targeted to cell wall components of MTB but possibly also and other surface molecules and secreted proteins | (131618252627303139) |
![]()
EMERGING THEME 1
Abs Associated with Latent Infection Are More Protective Than Those from TB Patients
New evidence for the role of Abs in TB can be found in a recent study by Lu et al. (9) who investigated the differences in Abs between patients with active TB disease and those with latent MTB infection. The study reported that individuals with latent and active TB have distinct Ab responses and that Abs associated with latent infection have superior functional properties. Importantly, pre-incubating MTB with latency-associated Abs before infection of human blood derived macrophages decreased the burden of infection significantly compared to pre-incubating MTB with Abs from active TB infection. This could be due to shuttling the bacilli through pathways resulting in intracellular killing via cell surface Fc receptors, or intracellular Fc receptors such as TRIM21 (10) as suggested by the authors.
Another study also set out to determine the effects of human anti-TB Abs but in a mouse model of infection. Thus, Li et al. (11) have investigated the protective role of polyclonal antisera from healthcare workers exposed to MTB. The study found that Abs (predominantly IgG3) from healthcare workers who were latently infected or uninfected showed a protective effect in mice challenged with MTB by the aerosol route. In agreement with the above studies, the Abs from healthcare workers were more protective than those from TB patients and this appears to be the most consistent emerging theme from these recent studies. However, it is unclear whether these human Abs could display their full protective potential in mice but the authors rely on at least partial cross-reactivity of the human-mouse Ig-FcR system (12).
Another recent study by Zimmermann et al. (13) reported generation of several human recombinant IgA and IgG anti-TB Abs using cells from TB patients and their exposed contacts. These Abs were then tested for their MTB inhibitory activity using the human lung epithelial cell line A549 and the primary human lung epithelial cells. In agreement with above 2 studies, it appeared that Abs from household contacts (presumably latently infected) performed better than Abs from TB patients. The limitation of this and the Lu et al. (9) study is that the MTB inhibition was performed only in vitro thus potentially missing on the Ab effects and mechanisms that are operational only in a complex in vivo environment.
EMERGING THEME 2
Glycosylation Profile Is Associated with Divergent Functions
Lu et al. (9) analysed the differences in glycosylation profiles of Abs between active TB and latent TB infection (LTBI) by capillary electrophoresis. Surprisingly, the 2 sets of Abs clustered as distinct glycan signatures although significant within-the-group heterogeneity was also observed, likely representing different spectra of the infection. Interestingly, serum IgG from LTBI individuals contained less fucose, which is known to enhance binding to FcγRIII (14) and higher galactose and sialic acid content, both of which are known to be anti-inflammatory. Similar trend was observed with both total and purified protein derivative-specific serum IgG indicating that active MTB infection may be capable of subverting the hosts Ab repertoire as a whole, just as it does with many other components of immune response. Indeed, in another recent study (15) it was reported that infection of mice with MTB modified glycosylation of serum proteins and notably IgM Abs and that fucosylation and sialylation were most affected.
These glycosylation differences could lead to new biomarkers of disease and also diagnostic tests to differentiate between active and LTBI. In the context of immunotherapy, the differences in glycosylation between active or LTBI and their potential influence in intracellular killing is of considerable interest as they may aid in the development of more efficacious monoclonal Abs
EMERGING THEME 3
Ab Isotype Plays a Role
Zimmermann et al. (13) tested several human recombinant IgA and IgG anti-TB Abs for their MTB inhibitory activity using human lung epithelial cells. Interestingly, the IgA Abs specific for lipoarabinomannan (LAM) and heparin-binding haemagglutinin (HBHA) restricted infection whereas IgG Abs exacerbated it. However, the isotype dependent contrasting activities of IgA and IgG observed in that study are somewhat puzzling considering that epithelial lung cells do not express the conventional Fc-Ig receptors although they do express the FcRn (neonatal) receptor. Unfortunately, the authors in this study reported only a limited evidence for the activity of these Abs in human macrophage cells which are known to express abundant Fc-γ and Fc-α receptors.
Nevertheless, the Zimmerman et al. (13) study is agreement with our own studies with murine (1617) and human — (18) and unpublished—monoclonal Abs that showed IgA to be superior to IgG Abs of the same Ag specificity, at least when applied mucosally.
Thus, our own early studies using monoclonal IgA and IgG Abs against α-crystallin (Acr) Ag of MTB demonstrated superior effect of IgA, as measured by reduced bacterial counts and granulomas in mice (1617). Furthermore, the IgA mAb was also shown to suppress relapse of infection after incomplete drug treatment (19). In both instances the IgA mAb was given to mice intranasally and the best treatment outcome was achieved when combining with IFN-γ, a proinflammatory cytokine which is thought to play an essential role in controlling MTB infection. We then generated a novel fully humanised IgA mAb (2E9IgA) (18) by screening a naïve human single chain fragment phagemid library against the MTB Acr Ag, an abundant Ag found on the surface and in the cytosol of MTB as well as BCG. Acr is upregulated in mycobacterial dormancy (2021), and this provides the basis of the rationale behind development of an anti-Acr mAb. To investigate the efficacy of 2E9IgA, we used BALB/c transgenic mice (2223), which express the CD89 receptor required for IgA binding to host cells. This was because wild type BALB/c mice do not express a homologue for CD89 (24), which is required for host mediated effects of IgA. CD89 transgenic mice were inoculated with 2E9IgA and IFN-γ (combined immunotherapy) intranasally and challenged with H37Rv either before or after combined immunotherapy. The level of infection was then quantified in lung and spleen homogenates, and we found that the combined immunotherapy significantly reduced colony-forming unit (CFU) in mice compared to the negative control group receiving saline solution (18).
Nevertheless, isotype superiority may also depend on Ag specificity since other studies have demonstrated that IgG Abs can also be protective (25–27). Thus, in early studies, Teitelbaum et al. (27) investigated 2 murine monoclonal Abs against arabinomannan (AM), 9d8 and 5c11, which were IgG3 and IgM, respectively (27). The study found that while both 9d8 and 5c11 showed binding to AM by ELISA, only 9d8 showed protection against in vivo challenge with virulent MTB in mice. Mice given 9d8 displayed increased survival compared to those treated with an isotype control, although their lung CFU counts remained unchanged. Interestingly, mice treated with 9d8 presented significantly higher granuloma formation, which the authors attributed to the release of cytokines such as TNF-α due to host mediated Ab responses.
In another study, however, an anti-LAM IgG1 given to mice intravenously reduced lung bacterial counts and granulomatous infiltration (26), demonstrating that anti-LAM IgG Abs can also be protective. In agreement with this, de Vallière et al. (28) demonstrated the role of LAM specific IgG Abs in human volunteers. Volunteers were given 2 doses of intradermal BCG 6 months apart, and serum samples containing MTB-specific IgG Abs were collected and shown to be predominantly directed to LAM. These Abs were tested in vitro where it was found that they increased internalization of BCG in phagocytic cells, suggesting that opsonisation is a potential mechanism of action for these Abs. The study also demonstrated increased killing of BCG by phagocytic cells in vitro after the addition of serum LAM Abs, possibly by increased phagosome-lysosome fusion, and this was specific to the IgG present in the serum sample as serum treated with protein G (to remove IgG) did not show increased killing of BCG.
Furthermore, reduction in post-chemotherapy relapse was reported in a study in which IgG Abs from mice immunised with detoxified MTB extract were transferred to naïve recipient mice (29). In addition to these studies, Abs of the IgG2a or IgG3 subtype against the HBHA reduced the infection in the spleens but not lungs of infected mice, suggesting that these particular IgG Abs could inhibit bacterial dissemination from lungs to other organs (30).
EMERGING THEME 4
Abs Target Surface and Nonsurface Ags
In a recent study, Prados-Rosales et al. (31) reported that a conjugate vaccine made of AM and Ag85B induced strong Ab responses to both components and when these Abs were transferred to naïve mice they were found to reduce MTB infection in a dose dependent manner. Interestingly, while the authors focus predominantly on AM-specific Abs, the fact that Ag8B also induced protective Abs is of utmost interest. Ag85B is not a classical target in anti-TB Ab studies as it is not a cell wall component but an enzyme involved in cell wall synthesis and then secreted into extracellular environment by the bacterium. While numerous studies (32–37) have shown that Ag85B is strongly immunogenic and potentially protective as a vaccine component, little is known about its role once secreted by the bacteria. If indeed Abs against this Ag are protective as evidenced in the Prados-Rosales et al. (15) study, this could mean that Ag85B has a secondary, virulence-associated role apart from being mycolyl-transferase involved in cell wall synthesis. Indeed, Ag85B can be found in abundance in lung cell homogenates of MTB infected mice (38) and could be involved in modulating bacterial environment to facilitate infection of the new host cells.
Another study also focused on AM Abs but this time induced by BCG vaccination (39). In that study, sera from BCG vaccinated subjects were tested for their opsonic properties and effect on phagocytosis and intracellular survival in human macrophages. The authors report enhanced phagocytosis and phago-lysosomal fusion when BCG was opsonised with Abs and this also resulted in restricted intracellular survival. The obvious limitation of this study is that BCG was used rather than MTB, and with the latter well known to be able to inhibit phago-lysosome fusion (4041), it remains to be seen if these effects could be reproduced with MTB.
While Ag85B represents an interesting target for anti-TB Ab action, it is not the only non-cell wall component that appears to be able to induce potentially protective Ab responses. While most of the studies still focus on classical cell wall components such as AM, LAM, or mannosylated LAM (262739), there are several other Ags that are currently of interest as Ab targets, including Ag85B (1331), early secreted antigenic target of 6 kDa (ESAT-6) (13), HBHA (16), Acr (1618), MPB83 (25) and a number of unidentified bacterial lysate Ags (13). Further antigenic targets for Abs have been identified in a recent study by Coppola et al. (42) who analysed IgG levels and specificities in sera of patients and controls in an endemic TB setting. They observed that Ab levels to particular Ags associated with bacterial burden and control vs progression of disease. Some of these Ags may appear on the cell surface transiently or indeed may be secreted to the cell culture, such as Ag85B and ESAT-6. Either way, they offer additional opportunities for Abs to exert their protective functions and so, the list of the potential new targets is constantly expanding. It is therefore quite plausible that some of these Abs could work synergistically, by targeting different bacterial components and virulence mechanisms employed at different stages of the infection.
POSSIBLE MECHANISMS OF Abs ACTION IN TB
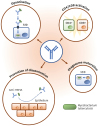
The protective mechanisms of Abs in TB are likely to be diverse and dependent on their Ag specificity, isotype and site of secretion or route of delivery if applied as immunotherapy. It is indisputable that the tubercle bacilli are released from necrotic lesions in significant numbers (43) and that means that Abs could gain access to these extracellular bacilli and influence both their FcR-mediated uptake by uninfected macrophages (182739) or dissemination throughout the lungs and other organs (30). Similar, but Fc-receptor independent action has been proposed also for mAb blocking of the action of LAM (26) by diverting the MTB away from the mannose receptors towards more bactericidal receptors. This may lead to enhanced phagocytosis, phago-lysosomal fusion and decreased intracellular survival, as proposed in some studies (182839). In addition, some IgG Abs were shown to enhance the mycobacterium specific IFN-γ production by CD4+ and CD8+ T cells (28) offering yet another potential mechanism of Ab mediated anti-TB immunity. Also, Abs may contribute to the maturation of dendritic cells and enhancement of MHC class I restricted presentation of antigenic peptides to T cells (44). Some of these potential mechanisms of Ab action against TB are schematically illustrated in Fig.1 but this is certainly not an exhaustive list and other yet unidentified mechanisms may come to light as the research in this field gathers pace.
THE EVIDENCE AGAINST
While all providing a supporting evidence that Abs can be protective in TB, none of the above studies has yet provided definitive, hard-core evidence that we called upon at the beginning of this review. It is therefore important to also consider the evidence that discounts the role of Abs in TB so that a balanced view can emerge. Thus, the strongest argument that Abs do not contribute to protection against TB comes from numerous preclinical vaccine studies showing strong Ab responses but minimal or no protection; an example of one such study is described in (45). In other words, Ab responses do not appear to be correlated with protection. However, as a counter argument to that, one can draw a parallel to IFN-γ, an important pro-inflammatory macrophage-activating cytokine in immunity to TB. Although the essential role of IFN-γ in TB is indisputable as shown by many experimental and clinical studies (46), it appears to be no more than a prerequisite of protection rather than a correlate of it. Similarly, Abs could have a significant role in protection but alone are not sufficient in the absence of other yet unidentified correlates of protection. This is quite plausible given that MTB infection is incredibly complex and stage driven so that different elements of immunity are required at different stages of infection to achieve control.
Another argument against the role of Abs comes from the B cell knockout mouse studies. One such recent study reported that B cells (and therefore Abs) are not required for protection against MTB imparted by a viral subunit mucosal vaccine (47). In contrast, T cells were shown to be essential for protection in that study. Again, a counter argument to that comes from previous studies showing contrasting results, where B cell deficient mice were shown to be more susceptible to MTB infection (48), while lung granulomas of non-human primates infected with MTB showed the presence of activated B cells (49). Furthermore, and as a yet another layer of complexity in regards to B cells, they were also proposed to have a regulatory role in TB (50).
CONCLUDING REMARKS
Further essential research is required in both animal experimental models and humans that could shed further light on the role of Abs in TB. Concerns may be raised about the impact of vaccine induced Abs in experimental models of infection as well as the efficacy of passive Ab treatment. The experimental results from the recent studies described in this review showed merely a reduction in counts of viable organisms, whether in mice or infected cell lines. This is in stark contrast to sterilising potency which can be imparted by drugs. This could be due to inadequate experimental models, high infection doses compared to humans and absence of other elements of immunity. Since no vaccine candidate against TB has yet imparted sterilising immunity in any of the experimental models, it is unlikely that Abs could achieve that alone—that would be setting the bar too high. However, the present encouraging evidence suggests they could yet play a significant role in both vaccine design and immunotherapy of TB. That paradigm shift has already occurred and it is only a matter of time before we can begin to harness the wide range of anti-mycobacterial activities. To that effect, the research over the past 5 years has given us some clear pointers and although the definitive evidence is yet to come, there are already several emerging themes. These themes represent the pieces of a jigsaw that is becoming more and more complete with each new piece of evidence in support of the protective role of Abs in TB.
 XML Download
XML Download