

 PDF
PDF Citation
Citation Print
Print



Abbreviations
cAMP
cyclic AMP
CAR
chimeric Ag receptor
DC
dendritic cell
FoxP3
fork head box P3
HIF-1α
hypoxia-inducible factor 1-alpha
MDSC
myeloid-derived suppressor cell
MSC
mesenchymal stem cell
NSCLC
non-small cell lung cancer
PGE2
prostaglandin E2
PI3K
phosphatidylinositol 3-kinase
PKA
protein kinase A
PLC
phospholipase C
RCC
renal cell carcinoma
siRNA
short interfering RNA
TAM
tumor-associated macrophage
TDMMC
terminally differentiated myeloid mononuclear cell
Th1
Th type 1
INTRODUCTION
Both chemotherapy and radiotherapy continue to be used as routine cancer treatment protocols, even though cancer is considered the second leading cause of death worldwide (1). Immunotherapy is a new approach and potential treatment for various cancers. Immunotherapeutic strategies aim to inhibit tumors by suppressing tumor tolerance mechanisms and augmenting the immune response that enable both identify and destruction of cancerous cells. Although the first series of cancer vaccines were shown to be safe and significantly induced specific immune responses, these vaccines were unable to provide effective results in the clinic (234). Tumors employ several mechanisms to escape immune responses by producing a variety of tumor-derived factors that could exert immunosuppressive effects to promote both invasion and metastasis (5).
Clinical trials recently changed to combination protocols of a tumor vaccine with other protocols such as anti-cancer drugs or radiation therapy. The rationale for this approach is that radiotherapy or chemotherapy may help tumor vaccines exert their desired effects. This approach aligns with the concept of conventional anti-cancer therapies and may be in accordance with therapeutically appropriate antitumor immune responses (67).
In 1863, Virchow was the first to discuss the relationship between inflammation that is promoted by cancerous tissue and tumor genesis (8). Inflammatory compounds contain reactive oxygen species and nitrogen intermediates, prostaglandins, growth factors, inflammatory cytokines, chemokines, and adenosine. All of these inflammatory entities are involved in tumor development by stimulating different mutations, resisting apoptosis, inducing angiogenesis, and facilitating a survival benefit to a sensitive cell (89).
In a cancer response, immune cells have binary actions with the potential to either remove or induce malignancy (1011). Cancer cells use the help of immune cells, like tumor-associated macrophages (TAMs), NKT cells, Tregs, and myeloid-derived suppressor cells (MDSCs) to generate locally in tumor-bearing hosts. Treg-mediated immunosuppression is both one of the principal tumor immune-evasion mechanisms and a crucial barrier to successful tumor immunotherapy (111213). MDSCs are a population of myeloid cell progenitors that differentiated to macrophages, dendritic cells (DCs), or other granulocytes. These MDSCs execute suppressive functions on T cell responses through nitric oxide, reactive oxygen species, prostaglandin E2 (PGE2), adenosine, and TGF-β production while also inducing Treg and anti-inflammatory responses (1415). It has become apparent that efficient cancer treatments need a different application, which mainly targets multiple pro-tumorigenic pathways and disrupts immunosuppressive networks (2).
Adenosine is a purine nucleoside with different well-known functions in various pathophysiological processes that is released from cells or generated extracellular (716). Adenosine can prevent immune cells involved in anti-tumor responses and stimulate the development of immunosuppressive cells such as Treg and MDSCs by adenosine receptors binding (1718). Adenosine and its receptors attracted researchers' attention in cancer pathology studies and create new research area in therapy purposes (1920). Here, we discuss both the pharmacologic and immunologic inhibition of adenosine metabolism, which along with other immunotherapies, can induce effective anti-tumor responses.
MOLECULAR BIOLOGY OF ADENOSINE
There are low levels of adenosine in unstressed tissues under physiological conditions, but they can rapidly rise in response to hypoxia-ischemia, inflammation, or trauma (21). When adenosine is released from intracellular sources into the extracellular space, it acts as an ‘alarm’ or danger signal. In addition, through its effects on cell surface receptors, it triggers numerous cellular responses that target tissue homeostatic mechanisms (22).
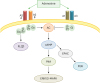
Adenosine is produced by the ectoenzyme CD73 from AMP, which is generated by the ectoenzyme CD39 from ATP. Adenosine fulfills its biological functions via four subtypes of adenosine receptors (A1, A2A, A2B, and A3), which belong to the G-protein-coupled family of receptors (23).
Adenosine receptors' mechanisms of action are inhibition or stimulation of adenylyl-cyclase to decrease or increase amounts of intracellular cyclic AMP (cAMP).Both A1 and A3 receptors decrease cAMP levels, whereas A2A and A2B increase cAMP contents. Adenosine receptors activate MAPK pathways and, in some cells, A1, A3, and A2B receptors direct phosphatidylinositol 3-kinase (PI3K) or Gq/phospholipase C (PLC) pathways (Fig. 1). The biological activity of adenosine is regulated by an ecto-adenosine deaminase, which converts adenosine into inosine molecules. Between these adenosine receptors, the A2A receptor plays a definitive role in the regulation of inflammatory responses. Different studies presented evidence about the activity of A2A receptor agonists as both anti-inflammatory and immunosuppressive mediators (24).
EFFECTS OF ADENOSINE ON THE IMMUNE SYSTEM
T cells
Extracellular adenosine via the A2A receptor can inhibit both activation and proliferation of CD4+ T cells and has been proposed not only to prevent Th type 1 (Th1) responses in vivo, but also, to prepare induction of Treg (25). T cells cultured in the presence of an A2A receptor agonist could neither proliferate nor generate IL-2, TNF-α, and IFN-γ. Other immunosuppressive roles of adenosine A2A receptor antagonists include the up regulation of T-cell negative regulatory molecules such as CTLA4, PD-1, and down regulation of the co-stimulatory molecule CD-40L. Stimulation of A2A with agonist induced T-cell tolerance and promoted Tregs and lymphocyte-activation gene 3 expression (26).
Tregs
A2A receptor stimulation enhanced fork head box P3 (FoxP3) mRNA and the immunosuppressor function of CD4+ FoxP3+cells. Since Tregs selectively co-express both CD39 and CD73, they may represent new specific markers of Tregs. Recent studies examined the relationship between the transcription factor FOXP3 and CD39, and found that FOXP3 up regulates the expression of CD39 (27).
A2A receptor stimulation in mixed-lymphocyte cultures raised the number of CD4+ FoxP3+ cell populations with superior immunoregulatory potency, while activation of effector T cells was mainly reduced so that this mechanism may be in accordance with the addition of Tregs within the tumor's microenvironment (2829).
Recently, Di Gennaro et al. (30) established that Treg isolated from different sites (peripheral blood, lymph nodes, and tumor infiltrating lymphocytes) of melanoma patients in stage III–IV, mediated immunosuppression by adenosine molecule. This study divided Tregs subsets with different CD39/CD73 expression into CD39+CD73− natural Tregs, CD39+CD73+ induced Tregs, CD39−CD73+ Tregs, and CD39−CD73– Tregs .The CD39−CD73− subset is unable to generate adenosine and its immunoregulatory activation might be exerted by secretion of TGF-β and/or IL-10, or cell-contact mechanisms (30).
DCs
DCs are professional Ag-presenting cells that up take Ags then process and present them to T cells. Therefore, DCs act as a connection between the innate and the adaptive immune systems (31). Activation of the A2A receptor in mature DCs leads to diminished IL-12, IL-6, and IFN-α but increases IL-10 secretion, so that it switches their cytokine pattern from a pro-inflammatory to an anti-inflammatory type (32).Adenosine actually controls TNF-β and IL-12 production; whereas it intensifies the production of IL-10 from LPS matured DCs. Adenosine matured DCs had a weakened ability to induce the differentiation of Th1 from naïve CD4+ T lymphocytes (33). DCs that generated in the presence of adenosine have altered function and produce high amounts of pro-inflammatory, angiogenic, immune suppressor, and tolerogenic components including IL-6, IL-8, and IL-10, cyclooxygenase-2, VEGF, TGF-β, and indoleamine-2, 3-dioxygenase; and promote tumor growth, if given to animal models (34). N-ethylcarboxamidoadenosine (adenosine analog) and forskolin (cAMP elevating-agent) skewed DCs differentiation to tolerogenic or regulatory subsets, which produced different immunosuppressor materials, and expressed myeloid/monocytic lineage markers. These types of DCs could not prime Ag-specific responses by CD8+ T-cells (35).
MDSCs
Cells with immunosuppressive properties have been stabilized and are called MDSCs. In mice; these cells display a population of myeloid cell lineages with both monocytic and granulocytic morphology and are usually distinguished by CD11b and Gr-1 markers (36). CD11b+Gr-1+ cells accumulate in the tumor microenvironment and lymphoid organs and are considered major contributors to tumor immunotolerance (37).
This observation indicates that MDSCs express high levels of the CD73 molecule, which is linked to high amounts of ecto-5′-nucleotidase enzymatic activity. This result suggested that targeting CD73 might reduce the population of MDSCs within the tumor medium and overcome their inhibitory condition (38). Ryzhov et al. (39) indicated that TGF-β signaling could induce the maturation of tumor-infiltrated MDSCs into terminally differentiated myeloid mononuclear cells (TDMMCs). These cells presented high levels of cell surface CD39 and CD73 and had a capacity of adenosine production. Also, deletion of the TGF-β receptor reduced TDMMCs' numbers, tumor growth rate, and metastasis (39). A study has shown that adenosine promotes MDSC function with the activation of A2B receptors, administration of A2B antagonist inhibited the accumulation of tumor-infiltrating MDSCs and enhanced Th1-like responses in the tumor environment and significantly delayed tumor growth (17). It was also demonstrated that A2B blockade raised the number of CD11b+Gr1+ cells in tumors and exclusion of MDSCs in mice significantly decreased VEGF production especially through A2B receptors (40).
NK cells
Adenosine prevents the capability of activated NK cells to destroy tumor cells and Perforin- and FasL-mediated cytotoxicity (41). Also, it was demonstrated that adenosine could be a main factor in blocking the ability of lymphokine-activated killer cells to kill tumor cells (42). Beavis et al. (43) suggested that the A2A antagonist has an anti-metastatic effect because it enhanced NK cell function. Hatfield and colleagues (44) established that hyperoxic therapy has an anti-metastatic impact, which was related to the hypoxia-adenosinergic immunosuppression of NK cells. A subset of NK cell peripheral blood CD16−CD56bright NK cells, produce high amounts of CD38-mediated adenosine and inhibit CD4+ T cell proliferation, and act as regulatory cells (45). NKT cells are a population of T lymphocytes, which are presented NK-associated molecules, such as NK1.1 and TCR on the surface. Like Treg, NKT cells express both ecto-nucleotidases CD39 and CD73, and expression of both enzymes plays a role in the regulatory functions of these cells. In addition, a subset of NKT cells termed invariant NKT cells requires adenosine A2A activation to generate both IL-4 and IL-10 (46).
Macrophages
The stimulation of adenosine receptors affects cytokine production by macrophages; it shows that the adenosine pathway mediates inhibition of TNF-α release mainly via A2A and A2B receptors (47). Activation of A2A receptors primarily has multiple inhibitory effects on the M1 macrophage subset, while adenosine receptors induce the M2 macrophage subset by up regulating the expression of several markers such as arginase 1, tissue inhibitor of matrix metalloproteinase 1, and macrophage galactose-type C lectin 1 (48). Several lines of evidence also support this idea that adenosine can increase VEGF secretion by macrophages through the activation of A2A receptors (49). Cekic et al. (50) investigated A2A expression on myeloid cells, specifically TAMs, and found indirectly mediated suppression of T cells and NK cell in the tumor microenvironment.
Neutrophils
It has been shown that stimulation of neutrophils by adenosine prevented attachment to vascular endothelial cells and damage to the endothelium by these cells (51). In addition, other studies found that the mechanisms of this inhibition by adenosine were through both selectin and integrin molecules (5253). Adenosine decreased secretion of oxygen radicals and other inflammatory mediators by neutrophils. Also, adenosine via A2A receptors diminished both the phagocytosis and apoptosis of neutrophils. Results of investigation indicate that cAMP-protein kinase A (PKA)-independent pathways are responsible for neutrophil inhibition by adenosine A2A receptors (Fig. 2) (545556).
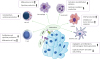
Figure 2
Inhibition of anti-tumor immune response. The purinergic pathway through adenosine molecules creates an immunosuppressive and tolerogenic condition in the tumor medium. The biologic function of different immune cells such as T cells, DCs, MDSCs, NK cells, macrophages, and neutrophils are influenced by adenosine, and aid in tumor progression.
ADENOSINE IN TUMORS ENVIROMENT
Extracellular amounts of adenosine in tumor tissues are higher than in normal tissues because of accumulation of ATP in the case of ischemia, damage, and stress. As mentioned previously, alteration of ATP to adenosine by the enzymatic activity of CD39 and CD73 in tumor tissues eventuates in immune response inhibition (5758). On the other hand, hypoxic media, which exist in the tumor medium, enhanced the breakdown of adenine nucleotides to adenosine (59). Binding of the transcription factor, hypoxia-inducible factor 1-alpha (HIF-1α), to the hypoxia response element containing the promoter region of the CD73 gene, induced transcription and protein expression (6061). Adenosine acting as an anti-inflammatory mediator down regulates the functions of infiltrating immune cells, thus preventing tissue destruction but this mechanism leads to tumor progression (62).
The expression of CD73 has been unregulated in many types of cancers, such as ovarian carcinoma (63), melanoma (64), prostate cancer (65), breast cancer (66), colon cancer (67), head and neck cancer (6869), leukemia (70), hepatocellular carcinoma (71), and glioblastoma (72). Many studies reported that CD73 expression in cancer is connected with several outcomes, such as progression, poor prognosis, metastasis, and weak response to chemotherapy agents (727374).
One of the new areas of interest in cancer research is MicroRNAs. Short non-coding RNAs bind to the 3′-untranslated regions of different genes and regulate their expression (75). Xie et al. demonstrated that miR-30a is a negative regulator of CD73 expression in colorectal cancer cells and leads to apoptosis and growth inhibition of tumor cells (76).
Different studies have shown that both expression and enzymatic activity of ecto-5′-nucleotidase (CD73) are elevated in various metastatic carcinomas (777879). Beavis et al. (43) explained that metastatic capability of CD73+ tumors is mediated by an NK cell-dependent mechanism. Recently, one study established that a BRA mutation is associated with increased CD73 expression in melanoma patients. In addition, adenosine enhanced lymph node pigmentation and promoted lymph node metastasis. Also, combining BRAF and A2A receptor inhibition decreased tumor growth rate and metastasis in experimental BRAF-mutant melanoma (80).
Previous studies found that if the activity of CD73 in cancer cells were inhibited, angiogenesis of the tumor would decrease, suggesting that this enzyme is involved in tumor angiogenesis (8182). Moreover, knockdown of CD73 expression in a breast tumor model diminished their metastatic potential for lung tissue (66).
Both CD39 and CD73 represent by the endothelial cells and can, therefore, be an origin of adenosine production. In addition, the presence of hypoxia in tumors increases the expression of both CD39 and CD73 on endothelial cells and leads to an increase in adenosine levels. The genetic or pharmacological deletion of CD39 and CD73 results in defects in the generation of tumor neovascularization (83). Further mechanistic investigations showed that adenosine can regulate the production of pro-and anti-angiogenic factors in tumor cells (8485). Indeed, adenosine induces the secretion of VEGF, IL-8, and angiopoietin 2 by several human cancer cell lines through A2B or A3 receptors (86). A2B receptor stimulation promotes the release of VEGF by host immune cells that infiltrated the tumor. Ryzhov et al (39). showed tumor-infiltrating CD45+ immune cells, by engaging A2B receptors, involved in VEGF production in a mouse lung carcinoma model. Deletion of Cd39/Entpd-1 in mice not only disrupted the angiogenesis process but also delayed the development of melanoma tumors. These data indicated that CD39/ENTPD-1 expression was associated with the formation of new vessels and tumor growth (87). The described pro-angiogenic roles of CD73 might be related to sustaining an immunosuppressive network in the tumor microenvironment.
Recent evidence proposed that mesenchymal stem cells (MSCs) in the tumor site play essential roles in the progression, invasion, metastasis, and induction of epithelial-to-mesenchymal transition of malignant cells. These cells are distinguished by 3 markers including CD73, CD105, and CD90; so, they might be a good source of adenosine generation (88). Although, the expression of CD39 on MSCs was not reported, other CD39+ cells like Treg can produce AMP in tumors (89). Also, some studies indicated that cancer derived MSCs can inhibit the function of antitumor-effective immune cells through adenosine production (9091).
ADENOSINE INHIBITION AND CANCER TREATMENT
Monoclonal Abs
Cancer treatment using monoclonal Abs has been demonstrated as one of the most successful therapeutic approaches in several tumors in the last 20 years (92).Several preclinical studies showed that pharmacological blockade of CD73, as well as treatment with anti-CD73 monoclonal Abs, is effective in preventing both tumor growth and metastatic spread in animal models (6572). One study argued that anti-human CD73 Abs, which targeted adhesion and catalytic activity of CD73, could inhibit the development of metastasis in a breast cancer model (93). The study by Allard et al. showed that blockade of CD73 with a monoclonal Ab significantly diminished tumor VEGF production, and prevented tumor angiogenesis in vivo (83). MEDI9447 is a human, high-affinity Ab that inhibits the hydrolysis of CD73 to AMP. An in vivo study showed that MEDI9447 could impair the suppression of immune effector cells by adenosine and lead to tumor growth inhibition. Moreover, blockade of CD73 with MEDI9447 resulted in elevated Ag presentation and enhanced lymphocyte activation; and therefore, led to higher production of inflammatory cytokines such as IFN-γ, IL-1β, and TNF by Th1 cells (94). Young et al. identified that targeting A2A receptor antagonism in association with an anti-CD73 Ab that employs Fcγ receptors, limited tumor development and metastasis. This study demonstrated that combined inhibition of CD73 and A2A receptor is more effective than inhibition of either alone (16).
Pharmacological inhibitors
Different adenosine receptor antagonists have been developed for numerous therapeutic applications, such as cardiovascular, inflammatory, and neurodegenerative diseases without any unwanted side effect (9596). Many studies showed that pharmacologic inhibition of adenosine especially through A2A and A2B, or CD73 and CD39 are clinically useful treatments in cancer (Table 1). Also, there are some studies about effect of A1 and A3 agonist on tumor development. It is established that specific agonist of A1 and A3 receptor could delay melanoma growth in CD73 knockout mice but increased angiogenesis (85).
Table 1
The effects of adenosine A2A and A2B receptors antagonist on animal cancer models

| Target | Drug | Tumor | Effect | Ref |
|---|---|---|---|---|
| A2AR | ZM241385, Caffeine | Lung | Enhanced antitumor effect of CD8+ T cells | (97) |
| Caffeine | Melanoma | Limitation of tumor neovascularization and increased apoptosis | (97) | |
| SCH58261 | Ovary | Prolong the survival of tumor-bearing mice | (63) | |
| Breast | Increase of doxorubicin activity against tumor cells | (73) | ||
| Melanoma | Inhibition of metastasis | (43) | ||
| Breast | Inhibition of metastasis | (43) | ||
| Melanoma | Inhibition of metastasis | (16) | ||
| Melanoma | Inhibition of tumor growth, induction tumor infiltration of NK and CD8+ cells | (80) | ||
| A2BR | ATL801 | Breast | Inhibition of tumor growth and metastasis | (98) |
| ATL801 | Bladder | Inhibition of tumor growth and Inducing T cell immune response | (98) | |
| PSB1115 | Melanoma | Inhibition of tumor growth and Inducing T cell immune response | (17) | |
| CVT-6883 | Lung | Decrease of VEGF and cAMP production | (99) | |
| CD73 | APCP | Breast | Inhibition of tumor migration | (43) |
| Melanoma | Enhanced tumor regression by production of Th1 cell-associated and Th17 cell-associated cytokines and CD8+ T cell infiltration in the tumor microenvironment. | (100) | ||
| Melanoma | Inhibition of tumor growth | (101) | ||
| Ovary | Increased survival of tumor-bearing mice | (63) | ||
| Melanoma | Inhibition of lung metastasis | (74) | ||
| Breast | Decrease of micro vessel formation in tumors | (83) | ||
| Breast | Inhibition of tumor growth | (102) | ||
| Melanoma | Inhibition of tumor growth | (103) | ||
| Melanoma | Inhibition of tumor growth and angiogenesis | (104) | ||
| Melanoma | Inhibition of tumor metastasis | (16) | ||
| Glioblastoma | Inhibition of tumor growth, migration and invasion | (105) |
Short interfering RNA (siRNA)
siRNA is used to regulate gene expression in various therapeutic approaches (106). Zhi and colleagues (82) demonstrated that blocking CD73 by siRNA suppressed CD73 gene and protein expression in the breast cancer cell line MB-MDA-231, leading to inhibition of both growth and metastasis inhibition in vivo. In another study by this group, CD73 siRNA delayed breast cancer growth both in vivo and in vitro by arresting the cell cycle in the synthesis phase and inhibited the apoptosis pathway (107). Jadidi-Niaragh et al. (108) designed CD73-siRNA encapsulated into chitosan-lactate nanoparticles, which were applied to inhibit CD73 molecules in an animal model of human metastatic breast cancer.
SIMULTANEOUS REMOVAL OF ADENOSINE AND CANCER IMMUNOTHERAPY
Because of the robust nature of the immune system such as its ability for memory and specificity, it is anticipated that cancer immunotherapy can achieve total, long-lasting remissions and cancer rejection with few or no side effects (109).
However, the presence of different factors with immunosuppressive capacity in the tumor microenvironment is a formidable obstacle in effective cancer immunotherapy. The presence of these factors indicated that immune regulatory cells such as Tregs, MDSCs, NKT cells, and TAMs are the important immunoregulatory cells that disrupt effective responses against tumors (9110).
Additionally, multiple soluble components such as HIF-1α, VEGF, and PGE2, inhibitory cytokines like IL-10 and TGF-β, and adenosine can also debilitate the efficacy of anti-tumor responses (9111). Therefore, the reduced amount of adenosine in the tumor medium may improve the effectiveness of cancer vaccine immunotherapy.
The progress in tumor biology regarding both the conception and potency of immune system-based cancer vaccines may derive from evidence demonstrating that genetic deletions of the A2A receptor or the blockade of A2A receptor signaling by A2A receptor antagonists both restored suppression of anti-tumor T cells and induced tumor rejection (97).
Components which target the A2A receptor pathway can induce antitumor immunity by limiting results of extracellular adenosine generated from tissues and Tregs. This observation provides considerable evidence for the high expression of both CD39 and CD73 ectoenzymes on Tregs, MDSCs, and MSCs that secrete adenosine and have various therapeutic applications (112).
T cell-based therapy and adenosine
T lymphocytes are the effector arms in the response to cancer and immunosurveillance. Accordingly, numerous therapeutic approaches have been generated to augment effector T cells against tumors (113). Ohta et al. (97) found that adoptively transferred CD8+ T cells in mice that received ZM241, 385 (A2A receptor antagonists) decreased metastasis in a CL8-1 melanoma model. In a study by Jin et al. (63) inhibition of the A2A adenosine receptor with the antagonist (SCH 58261and caffeine) rescued tumor-specific immune response and enhanced the efficacy of adoptive T-cell therapy. The combination of SCH58261 and adoptive T-cell therapy significantly could improve survival in mice compared with T-cell therapy or SCH58261 alone (63). Wang et al. (103) showed that a combination of T-cell immunotherapy with a CD73 inhibitor (APCP) inhibited tumor growth in a melanoma model compared with immunotherapy or APCP treatment alone. In this study, treatment with anti-CD73 and T-cell therapy in a peritoneal ovarian model increased survival of mice and was more effective than mono-therapy.
T lymphocytes, which were modified genetically to express a chimeric Ag receptor (CAR), have been successful in the treatment of some malignancies especially; hematologic cancers (114115). Beavis et al (116). showed that activation of the CAR unregulated expression of the A2A receptor. Also, genetic or pharmacological blockade of the A2A receptor increased CAR T-cell efficacy remarkably when associated with PD-1. This protocol leads to IFN-γ production of CD8+ CAR T-cells and induction of CD8+ and CD4+ CAR T-cells. This study showed that the A2A receptor pathway restricts the activity of patient-derived CAR T cells because CAR T cells that were also A2A receptor-deficient had a significantly better therapeutic influence than did the wild-type of CAR T (Fig. 3A) (116).
Figure 3
Efficacy of Immunotherapy. Inhibition of adenosine production and function, along with immunotherapeutic methods, may provide many advantages. DC vaccination and adenosine blockage could induce a tumor-specific T-cell response and increase efficacy of the vaccine. T cell or CAR T-cell immunotherapy in combination with CD73 inhibitor, or an A2 receptor antagonist could lead to tumor regression. An immune check point inhibitor, such as anti-PD-1, anti-CTLA-4, combined with adenosine inhibitors could tumor progression and metastasis.
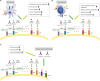
DC immunotherapy and adenosine
DCs are the primary target of different protocols of cancer immunotherapy because they can induce an effective and specific tumor immune response (117). In our study, we used an A2A adenosine receptor antagonist (SCH58261) and a APCP in combination with a DCs vaccine in a 4T1 breast cancer model. Combination therapy with SCH, APCP, and DC reduced tumor growth and VEGF production, improved survival of tumor-bearing mice, and induced specific antitumor immune responses (118).
Jadidi-Niaragh et al. (108) examined CD73-specific siRNA-loaded chitosan-lactate nano-particles with DC vaccine in 4T1 breast cancer model that led to reduced expression of CD73 on tumor cells, tumor growth, metastasis, and improved mice survival (119). Already, this nano-particle suppressed the expression of the CD73 molecule on tumor cells, in vitro (Fig. 3B).
As mentioned previously, activities of HIF-1α have a direct relationship to the adenosine pathway. The combination of HIF-1α inhibition and the DC vaccine also provided a cytotoxic T cell response, IFN-γ production, and increased efficacy of immunotherapy (120).
Immune check point blockade and adenosine
Immune check point blockades have good results in cancer treatment such as anti-CTLA-4 (ipilimumab) and anti-PD-1/PDL-1 (nivolumab, MK-3475/MPDL3280A, MDX-1105). Allard et al. (57) study can reveal that blockade of CD73 reinforced therapeutic activity of immune checkpoint inhibitors anti-CTLA-4 and anti-PD-1 monoclonal Abs. It proved that combination therapy in mice with the A2A receptor antagonist (SCH) and anti-PD-1 monoclonal Abs substantially reduced both experimental and spontaneous metastases, and prolonged the survival of mice when compared with monotherapy. This study suggested that inhibition of metastasis was dependent on NK, T CD8+, IFN-γ, and perforin (76).
Beavis et al. utilized co-blockade of PD-1 and adenosine receptor 2A in cancer treatment led to the significant expression of IFN-γ and Granzyme B by tumor-infiltrating CD8+ T cells, growth inhibition of CD73+ tumors, and extension of survival in mice. This study suggested that adenosine receptor 2A antagonists could improve the efficacy of anti-PD-1 monoclonal Abs in cancer therapy (77).
A recently published study has shown that combination therapy with anti-CD73 andanti-PD-1 improved survival, induced both an Ab-mediated response and an infiltration by T CD8+ cells in a murine ovarian cancer model (Fig. 3C) (4).
A number of clinical trials are investigating adenosine receptor antagonists or anti-CD73 and check point inhibitors. Novartis/Palobiofarma is testing a class of A2Areceptor antagonists PFB509 and PDR001 (anti-PD-1) in a Phase I trial in patients with advanced non-small cell lung cancer (NSCLC) (121). CPI-444, a small oral molecule, is an antagonist of the A2A receptor. Corvus Pharmaceuticals is studying single-agent therapy with CPI-444 for renal cell carcinoma (RCC) and in combination with atezolizumab (anti PD-L1 monoclonal Ab) for both RCC and NSCLC (122). Another study is in phase 1b with the combination of AZD4635 (A2A receptor antagonist) and durvalumab (anti-PD-L1 Ab) for NSCLC, metastatic castrate-resistant prostate carcinoma, and colorectal carcinoma (CRC) (123). In addition, combination therapy withMEDI9447 (anti-CD73) and MEDI4736 (human anti-PD-L1 IgG1 Ab) in advanced solid tumors, which are selected between adults, is being examined (124). Also, Corvus Pharmaceuticals designed a clinical trial to determine the efficacy of CD73 inhibitor (CPI-006) alone or in combination with an A2A receptor antagonist (CPI-444) and an anti-PD-1 Ab in several advanced solid tumors (Table 2) (125).
Table 2
Clinical trials with adenosine pathway inhibitors combined with an immune check inhibitor in cancer

| Target | Drug | Company | Study phase | Tumor | Combination agent | Code |
|---|---|---|---|---|---|---|
| A2AR | PFB509 | Novartis/Palobiofarma | I, Ib | NSCLC | PDR001 (anti-PD-1) | NCT02403193 |
| CPI-444 | Corvus Pharmaceutical | I, Ib | NSCLC, Melanoma, renal cell carcinoma, TNBC, colorectal cancer, bladder cancer | MPDL3280A atezolizumab (anti-PD-1) | NCT02655822 | |
| AZD4635 | AstraZeneca | I | NSCLC, metastatic castrate-resistant prostate carcinoma, colorectal cancer | MEDI4736, durvalumab (anti-PD-L1) | NCT02740985 | |
| PFB509 (NIR178) | Novartis | Ib | Solid tumors and non-Hodgkin lymphoma | PDR001 (anti-PD-1) | NCT03207867 | |
| CD73 | MEDI9447 | MedImmune | I | Selected solid tumor | MEDI4736, durvalumab (anti-PD-L1) | NCT02503774 |
| CPI-006 | Corvus Pharmaceutical | I | NSCLC, RCC, colorectal cancer, TNBC, cervical cancer, ovarian cancer, pancreatic cancer, endometrial cancer, sarcoma, SCC of head and neck, bladder cancer, metastatic castrate-resistant prostate carcinoma | CPI-004 (A2AR antagonist), pembrolizumab (anti-PD-1) | NCT03454451 |
CONCLUSION
Different treatments were defined by targeting important pathways or particular molecules that are essential for both tumor cell development and invasion. Many recent studies offer combination therapies that present suitable options in cancer immunotherapy for patients. Also, blocking any suppressor molecule or cell in the tumor microenvironment will increase the performance of different immunotherapeutic methods. The CD39/CD73 purinergic pathway can now be distinguished as one of the most substantial immunosuppressive regulatory mechanisms in the tumor medium. It is essential to try to develop approaches to reduce adenosine receptors and CD73 expression on both tumor and immune cells for subsequent adoptive immunotherapy. It seems that further studies are necessary to assess anti-tumor immune responses and design an effective targeted therapy to both disrupt and modulate the immunosuppressive network. These studies may suggest new strategic direction as well as provide insight for innovative approaches to overcome specific difficulties encountered in cancer therapy.
 XML Download
XML Download