

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Fluoroquinolones (FQs) are widely used to treat various bacterial infections [12]. FQ resistance has increased globally in Acinetobacter species, which are clinically important pathogens that frequently cause infections among intensive care unit patients [2345].
In A. baumannii, FQ resistance occurs mainly through mutations in the quinolone resistance-determining regions (QRDRs) of DNA gyrase (gyrA) and topoisomerase IV (parC), although overexpression of efflux pumps can contribute to FQ resistance [678910]. The most frequently described mutations in A. baumannii are within the QRDRs at Ser-83 in GyrA and at Ser-80 and Glu-84 in ParC [67811]. In particular, a double mutation, affecting the Ser-83 of GyrA and Ser-80 or Glu-84 of ParC, renders A. baumannii highly FQ resistant [711]. The single mutation affecting Glu-87 of GyrA, an important mutation associated with FQ resistance in other gram-negative microorganisms [12], has rarely been found in A. baumannii [8]. Detection of these mutations is therefore important for assessing FQ resistance in A. baumannii and epidemiological studies of resistant strains.
Although DNA sequencing is a reliable technique for detecting mutations, it is costly, time-consuming, and laborious when analyzing numerous clinical strains. As an alternative, PCR-restriction fragment length polymorphism (RFLP) has been used to detect mutations associated with FQ resistance in A. baumannii [6713]. However, this approach was insufficient for identifying significant mutations linked to high-level FQ resistance because it determined only the presence or absence of gyrA mutations at codon 83 or parC mutations at codon 80.
We have previously reported a mismatched PCR-RFLP assay for detecting gyrA and parC mutations associated with FQ resistance in Enterobacteriaceae [14]. We aimed to develop a mismatched PCR-RFLP assay to detect mutations in gyrA (codons 83 and 87) and in parC (codons 80 and 84) associated with FQ resistance in A. baumannii.
METHODS
Bacterial strains
We used 58 A. baumannii strains and 37 non-baumannii
Acinetobacter strains, including some of the strains used in previous studies [1516]. These strains were collected between 2009 to 2018 and stocked in our laboratory. The 37 non-baumannii
Acinetobacter strains included 10 A. nosocomialis, eight A. pittii, two A. calcoaceticus, two A. soli, two A. ursingii, two A. colistiniresistens, two A. johnsonii, two A. gerneri, one A. radioresistens, one A. bereziniae, one A. towneri, one A. grimontii, one A. junii, one A. tandoii, and one A. haemolyticus strain. These strains were identified by RNA polymerase β-subunit gene sequence analysis [17]. All strains were used to confirm the practicality of the mismatched PCR-RFLP assay. A. baumannii strain ATCC 19606 was used for comparison with the A. baumannii strains. According to the ethical guidelines for epidemiological studies released by the Ministry of Health, Labour, and Welfare in Japan [18], ethical approval and written or verbal informed consent are not required for this type of study.
Antimicrobial susceptibility testing
Susceptibility was tested using an agar dilution assay according to the Clinical and Laboratory Standards Institute (CLSI) guidelines [19]. The minimum inhibitory concentrations (MICs) of levofloxacin and ciprofloxacin were determined. To examine the effect of efflux pumps in FQ resistance, MICs were also measured in the presence of the efflux pump inhibitor carbonyl cyanide m-chlorophenylhydrazone (CCCP), which was incorporated into the Mueller–Hinton agar at a concentration of 12.5 µM [20].
Amplification and DNA sequencing of the gyrA and parC QRDRs
The sequence of A. baumannii strain ATCC 19606 was used as the reference susceptible strain. The gyrA and parC QRDRs from 58 A. baumannii strains were amplified using PCR and sequenced using Applied Biosystems 3730 DNA analyzer (Applied Biosystems, Foster City, CA, USA), as previously described [21]. The results were compared with those of the mismatched PCR-RFLP.
Development of the mismatched PCR-RFLP assay
Based on the conserved sequences of A. baumannii
gyrA and parC, we designed two sets of mismatched PCR-RFLP primers to detect mutations in gyrA (codons 83 and 87) and parC (codons 80 and 84) by introducing an artificial restriction enzyme cleavage site into the PCR products, as described previously [22]. The reverse primers for gyrA and parC are located immediately downstream of the nucleotide sequences, corresponding to GyrA87 and ParC84, with mismatched nucleotides to create recognition site (XmnI), respectively (Fig. 1). The primer sequences and PCR conditions were expected to yield 143 and 120 bp DNA fragments for gyrA and parC, respectively. We performed PCR amplification of gyrA or parC from each strain using the AmpliTaq Gold 360 Master Mix (Applied Biosystems, Foster City, CA, USA), according to the manufacturer's instructions and as described in Table 1. The gyrA and parC PCR products were digested with HinfI or XmnI at 37℃ for one hour, and the digested products were analyzed by electrophoresis using 3.0% MetaPhor Agarose gels (Takara, Shiga, Japan).
RESULTS
Susceptibility testing and DNA sequencing of the gyrA and parC QRDRs
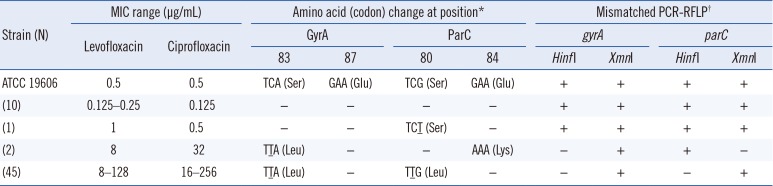
We analyzed 58 A. baumannii strains (47 FQ-resistant and 11 FQ-susceptible). Point mutations were present only in gyrA (codon 83) and parC (codons 80 and 84). The strains with no mutations (N=11) resulting in amino acid changes at Ser-83 and Glu-87 in GyrA and at Ser-80 and Glu-84 in ParC were susceptible to levofloxacin and ciprofloxacin. The FQ-resistant strains (N=47) carried two mutations, in gyrA (codon 83) and parC (codon 80 or 84), and all but one strain had ciprofloxacin MICs ≥32 µg/mL (Table 2). Of these 47 FQ-resistant strains, 45 possessed gyrA codon 83 TCA (Ser)→TT̲A (Leu) and parC codon 80 TCG (Ser)→TT̲G (Leu) mutations, and the remaining two possessed gyrA codon 83 TCA (Ser)→TT̲A (Leu) and parC codon 84 GAA (Glu)→A̲AA (Lys) mutations. CCCP did not appreciably affect the levofloxacin and ciprofloxacin MICs for the A. baumannii strains. The FQ-resistant strains were still classified as resistant based on the CLSI breakpoints for levofloxacin and ciprofloxacin, even in the presence of this efflux pump inhibitor. The mutations identified in gyrA and parC and the MICs of levofloxacin and ciprofloxacin are shown in Table 2.
Mismatched PCR-RFLP
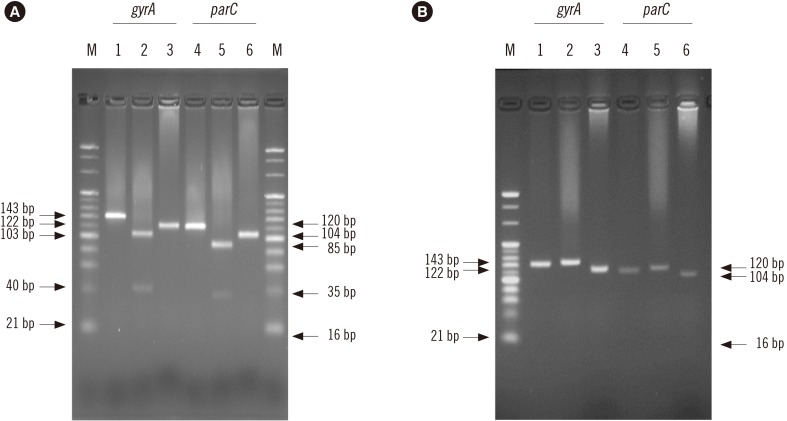
We performed PCR-RFLP on 58 A. baumannii strains and A. baumannii strain ATCC 19606. Amplification products with the expected sizes of 143 bp for gyrA and 120 bp for parC were successfully obtained for all strains. The PCR products of gyrA from FQ-susceptible strains contained both one natural HinfI recognition site and one artificially created XmnI recognition site. Similarly, the PCR products of parC from FQ-susceptible strains contained both one natural HinfI recognition site and one artificially created XmnI recognition site (Fig. 1). Consequently, HinfI and XmnI digested the amplified 143 bp fragment to generate two fragments of 103 and 40 bp and of 122 and 21 bp, respectively. The 21 bp fragment, which was produced by XmnI digestion, was not visible in Fig. 2; however, it was easy to recognize that the amplified 143 bp fragment was digested by XmnI. Similarly, HinfI and XmnI digested the amplified 120 bp fragment to generate two fragments of 85 and 35 bp and of 104 and 16 bp, respectively. The 16 bp fragment, produced by XmnI digestion, was not visible in Fig. 2; however, it was easy to recognize that the amplified 120 bp fragment was digested by XmnI. HinfI and XmnI failed to digest the PCR products at the site containing the mutations that resulted in amino acid changes at Ser-83 in GyrA or at Ser-80 or Glu-84 in ParC.
Furthermore, we applied this assay to 37 non-A. baumannii strains. While gyrA amplicons were obtained for 28 strains, namely 10 A. nosocomialis, eight A. pittii, two A. calcoaceticus, two A. ursingii, two A. gerneri, two A. johnsonii strains, one A. grimontii, and one A. tandoii, no parC amplicons were obtained in any of the 37 strains tested.
DISCUSSION
We developed a mismatched PCR-RFLP assay to detect mutations in gyrA (codons 83 and 87) and parC (codons 80 and 84), which are associated with FQ resistance in A. baumannii. This assay specifically detected significant mutations associated with reduced susceptibility to FQs in A. baumannii and accurately classified all the FQ-resistant and FQ-susceptible strains according to the MIC results.
The regions containing the mutation site resulting in amino acid change within the Ser-83 codon in GyrA or Ser 80 codon in ParC have a naturally occurring HinfI restriction site; thus, these regions are amplified by PCR, and the mutations at these positions are detected when the PCR products are not digested with HinfI, as analyzed by electrophoresis on agarose gels [67]. However, the mutation sites within the Glu-87 codon in GyrA and the Glu-84 codon in ParC are not involved at any restriction cleavage site. To detect mutations in gyrA (codon 87) and parC (codon 84), we introduced base substitutions near the mutation sites to create XmnI cleavage sites using the primer-specified restriction site modification method [22]. Our results demonstrated that while HinfI and XmnI digested the gyrA and parC amplicons from FQ-susceptible strains, they did not digest those from FQ-resistant strains with mutations in gyrA and parC. The mutations detected by this assay were concordant with the DNA sequencing results shown in Table 2. The high specificity of the restriction enzyme XmnI to each set of the three nucleotides for codon 87 (Glu) of GyrA and codon 84 (Glu) of ParC in FQ-susceptible strains allows our assay to accurately detect mutations at codon 87 in gyrA and at codon 84 in parC. Therefore, our assay can identify significant mutations in gyrA and parC QRDRs linked to high-level FQ resistance in A. baumannii without the need for DNA sequencing; this assay may thus serve as an alternative to other PCR-RFLP assays that are limited by their ability to detect mutations at only codon 83 of gyrA or codon 80 of parC [67].
Our assay accurately identified FQ-susceptible strains and FQ-resistant strains. When our assay did not detect mutations in gyrA (codons 83 and 87) and parC (codons 80 and 84), the MICs showed susceptibility to FQs. On the other hand, when our assay detected mutations in both gyrA and parC, the MICs showed FQ resistance. In addition, our assay enables simultaneous analysis of many strains and provides results within four hrs. Thus, it could aid in the rapid identification of FQ-resistant A. baumannii strains in the clinical setting.
Previous studies have reported several rapid assays for detecting gyrA and parC mutations associated with FQ resistance in A. baumannii and determining FQ resistance, including PCR followed by electrospray ionization mass spectrometry and pyrosequencing assay [2123]. These assays successfully detected FQ resistance mutations in gyrA (codon 83) and parC (codons 80 and 84) and identified FQ-resistant strains. However, these assays require expensive equipment; our assay does not.
Another important and advantageous finding was that the mismatched PCR-RFLP primers for parC amplified parC from A. baumannii strains, whereas these primers did not amplify parC from any non-baumannii
Acinetobacter strains. This assay may therefore differentiate A. baumannii from other Acinetobacter species in bacterial colonies, and determine FQ resistance for A. baumannii without the need for precise species identification within the genus Acinetobacter. This could also be an advantage of our assay. However, further studies are needed to confirm this finding.
The limitation of our assay is that it is unable to detect mutations at other locations of gyrA and parC or in other genes. Nevertheless, our data suggest this assay specifically amplifies gyrA and parC from A. baumannii and allows for simple, specific, rapid, and inexpensive detection of significant FQ resistance mutations. Thus, this assay may be useful for rapid assessment of FQ resistance in A. baumannii and for epidemiological studies of resistant strains in the clinical setting; moreover, it might be used to differentiate A. baumannii from other Acinetobacter species.

 XML Download
XML Download