

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Osteosarcoma is a malignant and rare tumor that primarily affects bones in juveniles and adults.12 It has been reported that osteosarcoma patients with metastatic or recurrent disease fare poorly, with overall survival rates of less than 20%.3 Therapeutic strategies, especially chemotherapy, lead to improved survival and outcomes in patients with osteosarcoma.456 Epirubicin is one of the most effective drugs against cancer and has gained widespread use in multiple cancers.78 Moreover, studies have shown that epirubicin could enhance the antitumor activity in osteosarcoma.910 Therefore, combination chemotherapy with epirubicin and other agents may provide an effective approach for osteosarcoma patients.
MicroRNAs (miRNAs), a class of small noncoding RNAs, can act as tumor-suppressors in tumorigenesis and development.11 Research has revealed that inhibition of miR-598 expression stimulates proliferation, migration, and invasion in osteosarcoma. Also, restoration of miR125a5p results in repressing osteosarcoma cell migration, invasion and epithelialmesenchymal transition by targeting matrix metallopeptidase-11.12 Moreover, it is well documented that miRNAs are involved in chemotherapy of cancers.1314 Li, et al.15 noted that a combination of miR-19a and miR-205 in the serum may predict chemosensitivity to epirubicin plus paclitaxel neoadjuvant chemotherapy for the luminal a subtype of breast cancer. Furthermore, miR-671-5p has been found to sensitize breast cancer cells to cisplatin, 5-fluorouracil, and epirubicin exposure and to reduce DNA repair capability in post-drug exposed breast cancer cells.16 Zhang, et al.17 found that miR-4443 depletion increases the sensitivity of epirubicin-resistant H1299 cells to epirubicin by decreasing the IC50 of epirubicin and inducing cell apoptosis and G0/G1 cell cycle arrest in non-small cell lung cancer. Additionally, previous studies have demonstrated that miRNAs are also correlated with chemosensitivity in osteosarcoma.18 For example, miR-125b was found to enhance chemosensitivity to cisplatin in osteosarcoma.19 miR-302b increased epirubicin-mediated anti-proliferation and pro-apoptosis effect in osteosarcoma.20 Recently, miR-1301 was reported to inhibit migration and invasion by targeting B-cell chronic lymphocytic leukemia/lymphoma 9 in osteosarcoma.21 However, whether miR-1301 is responsible for anti-osteosarcoma effects remains to be elucidated. TP53-regulated inhibitor of apoptosis 1 (TRIAP1) is overexpressed in many cancers and acts as an oncogene in tumorigenesis and development.22 Nevertheless, still relatively little is known about its role in osteosarcoma.
In this study, we hypothesized that TRIAP1 may be a target of miR-1301 and investigated the role of miR-1301/TRIAP1 in epirubicin-mediated anti-osteosarcoma effects.
MATERIALS AND METHODS
Cell culture and treatment
Human osteoblast hFOB1.19 cells and human osteosarcoma U2OS and SAOS-2 cells were purchased from American Type Culture Collection (Manassas, VA, USA) and cultured in the Dulbecco's Modified Eagle's Medium (Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% fetal bovine serum (Thermo Fisher Scientific) and 1% penicillin/streptomycin stock solution (Beyotime, Shanghai, China). Cells were maintained in a humidified atmosphere containing 5% CO2 at 37℃. U2OS and SAOS-2 cells (80% confluence in 6-well plates) were treated with different concentrations (0, 0.1, 0.25, 0.5, 1, 2.5, 5, or 10 µg/mL) of epirubicin (MedChemExpress, Princeton, NJ, USA) for 24 or 48 h. U2OS and SAOS-2 cells were transfected with miR-NC mimics, miR-1301 mimics, miR-NC inhibitors, miR-1301 inhibitors, pcDNA vector (vector), and TRIAP1 overexpression plasmid (TRIAP1). After 24-h transfection, cells were treated with epirubicin (1 µg/mL) for 48 h. The transfected or treated cells were used for subsequent experiments.
qRT-PCR assay
TRIzol (Beyotime) was introduced to extract total RNAs from cells according the manufacturer's instructions. Then, RNA was reversely transcribed into complementary DNA using TaqMan MicroRNA Reverse Transcription Kits (Biosystems, Foster City, CA, USA) or TIANScript RT Kits (Baolebo biotech, Beijing, China). Quantitative real-time polymerase chain reaction (qRT-PCR) analyses were carried out using Power SYBR Green kits (Takara, Shiga, Japan). The primers were as follows: TRIAP1 forward, 5′-AGGATTTCGCAAGTCCAGAA-3′, and reverse, 5′-GCTGATTCCACCCAAGTAT-3′; glyceraldehyde-3-phosphate dehydrogenase (GAPDH) forward, 5′-AGAAGGCTGGGGCTCATTTG-3′, and reverse, 5′-AGGGGCCATCCACAGTCTTC-3′; miR-1301 forward, 5′-ACACTCCAGCTGGGTTGCAGCTGCCTGGGAGTGA-3′, and reverse, 5′-CTCAACTGGTGTCGTGGA- 3′; U6 forward, 5′-CTTCGGCAGCACATATAC-3′ and reverse, 5′-GAACGCTTCACGAATTTGC-3′. The relative expression levels of miRNA (normalized to U6 small nuclear RNA) and mRNA (normalized to GAPDH) were evaluated by the 2-ΔΔCt method.
CCK-8 assay
Osteosarcoma cells (5000 cells/well) were seeded in 96-well plates and detected for cell proliferation using Cell Counting Kit-8 (CCK-8) assay according to the manufacturer's instructions. Briefly, the cells were incubated with 10 µL of CCK-8 (AbMole BioScience, Houston, TX, USA) at 37℃ for 4 h. Absorbance at 450 nm was evaluated using a microtiter plate reader (Thermo Fisher Scientific). Each group was tested with three replicates.
Cell apoptosis assay
Cell apoptotic rate was evaluated as described in a previous study using Annexin V-PE/7-AAD apoptosis assay (KeyGEN BioTECH, Nanjing, China).20 Briefly, cells were washed with cooled phosphate buffer saline and trypsinized. Annexin V-PE (1 µL) and 7-AAD (5 µL) were added in a dark room for 10 min incubation. Cell apoptotic rate was detected by a FACSCalibur flow cytometer with Cell Quest software (BD Biosciences, Franklin Lakes, NJ, USA).
Western blot assay
Cells were collected in Radio Immunoprecipitation Assay lysis buffer containing protease inhibitors (Beyotime). Cell lysates were centrifuged, and supernatants were collected and mixed with 6× loading buffer and boiled for 10 min. Proteins were quantified using bicinchoninic acid Protein Assay Kits (Solarbio, Beijing, China), separated with sodium dodecyl sulfate polyacrylamide gel, and transfected onto polyvinylidene fluoride membranes (Millipore, Billerica, MA, USA). The primary antibodies used in the present study were anti-Bcl-2: ab32124, anti-Bax: ab32503, anti-cleaved-caspase-3: ab2302, anti-cleaved-poly (ADP-ribose) polymerase-1 (anti-cleaved-PARP1): ab32064, anti-TRIAP1: ab225938, and anti-GAPDH: ab181602, which were purchased from Abcam (Cambridge, MA, USA). Horseradish peroxidase conjugated-secondary anti-body (goat anti-rabbit IgG: ab150077) was obtained from Cell Signaling Technology (Massachusetts, MA, USA). The protein signals were visualized using enhanced chemiluminescence (Thermo Fisher Scientific). Densitometry values were normalized to levels of GAPDH and analyzed using ImageJ 1.48v software (National Institutes of Health, Bethesda, MD, USA).
Luciferase reporter assay
The 3′-UTR of TRIAP1 containing predicted miR-1301 binding sites [wild-type TRIAP1 (WT) and mutant TRIAP1 (MUT)] were subcloned into the pmirGLO vector (Promega, Madison, WI, USA). Then, U2OS and SAOS-2 cells were cultured in 24-well plates and co-transfected with TRIAP1-WT or TRIAP1-MUT and miR-1301 mimics or miR-NC mimics, respectively. The relative luciferase activities were measured using the Dual-Luciferase Reporter Assay System (Promega).
Statistical analysis
All data are expressed as a mean±standard deviation. Statistical analysis was conducted using SPSS 18.0 software (SPSS, Inc., Chicago, IL, USA). Results were considered statistically significant only if the p value was less than 0.05 using Student's t test or one-way ANOVA. Our research was approved by Gansu Provincial Hospital.
RESULTS
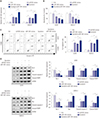
Effects of epirubicin on proliferation and apoptosis in osteosarcoma cells
First, to determine the effect of epirubicin on osteosarcoma cells, U2OS and SAOS-2 cells were treated with epirubicin at various concentrations. The results of CCK-8 assays showed that epirubicin inhibited osteosarcoma cell proliferation in a dose-dependent manner (Fig. 1A). Furthermore, 1 µg/mL of epirubicin was regarded as a standard concentration, since it provided effective inhibition of cell viability and a good amount of surviving cells. Then, the effect of epirubicin on cell apoptosis was further evaluated. We demonstrated that cell apoptotic rate was higher in cells with epirubicin exposure than that of cells treated with 0 µg/mL of epirubicin (Fig. 1B). Moreover, the detailed molecular mechanisms of epirubicin affecting apoptosis were subsequently explored. The level of anti-apoptosis protein Bcl-2 was decreased, while the expressions of Bax, cleaved-caspase-3 and cleaved-PARP1 were increased in U2OS and SAOS-2 cells treated with 1 µg/mL of epirubicin, compared with cells without epirubicin treatment (Fig. 1C), indicating that epirubicin participates in pro-apoptosis and anti-proliferation in osteosarcoma in vitro.
Epirubicin stimulates the expression of miR-1301 in U2OS and SAOS-2 cells
Then, we continued to examine the levels of miR-1301 in osteosarcoma cells. We observed that miR-1301 was downregulated in U2OS and SAOS-2 cells, compared with that of hFOB1.19 human osteoblast cells (Fig. 2A), suggesting that miR-1301 may act as a tumor suppressor in osteosarcoma. Moreover, whether epirubicin regulates the expression of miR-1301 was further addressed. As expected, epirubicin increased the abundance of miR-1301 in U2OS and SAOS-2 cells relative to that of the Ctrl group (Fig. 2B), demonstrating that miR-1301 may play a vital role in epirubicin-mediated effects in osteosarcoma cells.
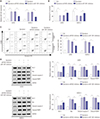
Upregulation of miR-1301 contributes to the anti-cancer effect of epirubicin in osteosarcoma
Considering that the level of miR-1301 was increased in osteosarcoma cells exposed to epirubicin, the role of miR-1301 in epirubicin-mediated anti-osteosarcoma was subsequently investigated. The results of transfection efficiency revealed that the introduction of miR-1301 mimics and epirubicin alone induced the enforced expression of miR-1301. Moreover, the promotion effect was enhanced by combination of epirubicin and miR-1301 mimics (Fig. 3A). As shown in Fig. 3B, the data further demonstrated that cell proliferation was impeded in the miR-1301 mimics group. More importantly, the inhibition effect was strengthened by a combination of them. Furthermore, the results of flow cytometry presented that cell apoptotic rate was elevated in cells transfected with miR-1301 mimics or treated with epirubicin. Intriguingly, a combination of miR-1301 overexpression and epirubicin reinforced the boosted effect on cell apoptosis (Fig. 3C). In addition, the expressions of apoptosis-related proteins were also analyzed in U2OS and SAOS-2 cells using Western blot. Our data showed that the combination of epirubicin and miR-1301 mimics increased the effects observed with separate treatment of them on the expressions of apoptosis-related protein (Fig. 3D), which was consistent with the data of flow cytometry. Consequently, our findings demonstrated that miR-1301 contributes to the epirubicin-mediated anti-osteosarcoma effect via a regulatory effect on cell proliferation and apoptosis.
MiR-1301 depletion weakens the anti-cancer effect of epirubicin in osteosarcoma
Based on the above results, we further conducted cell transfection to examine whether miR-1301 depletion weakens the anti-osteosarcoma effect of epirubicin. As displayed in Fig. 4A, the transfection of miR-1301 inhibitors abolished epirubicin-induced miR-1301 promotion. Simultaneously, cell proliferation and apoptosis continued to be evaluated. Downregulation of miR-1301 abated the epirubicin-mediated inhibition of cell proliferation in U2OS and SAOS-2 cells (Fig. 4B). Moreover, epirubicin promoted cell apoptosis, which was undermined by downregulation of miR-1301 (Fig. 4C). To expound on this observation, Western blot analysis of apoptosis-related proteins was also performed (Fig. 4D). The data further verified that miR-1301 knockdown overturned the pro-apoptosis effect of epirubicin in osteosarcoma cells.
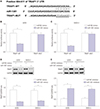
TRIAP1 is a target of miR-1301
In order to explore the molecular mechanisms of miR-1301 involvement in epirubicin-mediated anti-osteosarcoma effects, we used the TargetScan online database to search for potential candidate targets of miR-1301 and found that TRIAP1 contained binding sites with miR-1301 (Fig. 5A). Moreover, luciferase activity was dramatically reduced in the cells overexpressing miR-1301 in the TRIAP1 WT group, while it showed no significant change in the TRIAP1-MUT group (Fig. 5B and C), suggesting that TRIAP1 is a target of miR-1301. Furthermore, we also determined the effect of miR-1301 on expression of TRIAP1. The results showed a distinct suppression of TRIAP1 in miR-1301-upregulated cells, whereas inhibition of miR-1301 boosted it in U2OS and SAOS-2 cells (Fig. 5D and E), indicating that miR-1301 negatively regulates the expression of TRIAP1.
Epirubicin regulates TRIAP1 expression by modulating miR-1301
Finally, we also studied whether miR-1301/TRIAP1 axis is associated with epirubicin-mediated anti-osteosarcoma effects. qRT-PCR was subsequently employed to determine the mRNA levels of TRIAP1. We found that the mRNA levels of TRIAP1 were remarkably lower in epirubicin-treated osteosarcoma cells, which was attenuated by transfection of miR-1301 inhibitors (Fig. 6A). Similarly, Western blot also demonstrated that epirubicin inhibited protein levels of TRIAP1 in U2OS and SAOS-2 cells, which was rescued by miR-1301 knockdown (Fig. 6B and C). Subsequently, rescue-of-function was also performed to determine the effect of TRIAP1 upregulation on epirubicin-mediated TRIAP1 inhibition. Our data further indicated that overexpression of TRIAP1 abated lower expression of TRIAP1 induced by epirubicin at mRNA (Fig. 6D) and protein (Fig. 6E and F) levels. These data revealed that epirubicin repressed TRIAP1 expression by upregulating miR-1301. In addition, ectopic expression of TRIAP1 rescued the epirubicin-mediated decrease of cell viability (Fig. 6G). These results supported the hypothesis that miR-1301/TRIAP1 may be implicated in epirubicin-mediated pro-apoptosis and anti-proliferation in osteosarcoma.
DISCUSSION
Epirubicin, an anti-tumor drug, has been used in the treatment of osteosarcoma.20 In the current study, we found that epirubicin curbed cell proliferation and induced apoptosis in osteosarcoma cells, which was in line with a previous study.20 Emerging evidence suggests that miR-1301 could serve as a tumor-suppressor in a variety of cancers. For example, Zhi, et al.23 disclosed that miR-1301 is downregulated in glioma tissues and cell lines and inhibits cell proliferation by targeting neuroblastoma ras viral oncogene homolog. miR-1301-3p is decreased in breast cancer tissues and cells, induces cell cycle G0/G1 phase arrest and apoptosis, and inhibits cell proliferation by targeting immature colon carcinoma transcript-1.24 Here, we demonstrated that miR-1301 is expressed at low levels in osteosarcoma cells, suggesting that miR-1301 might contribute to the anti-osteosarcoma effect. Interestingly, the expression of miR-1301 showed an evident increase in cells after epirubicin exposure. These data indicated that miR-1301 may play an important role in osteosarcoma therapy in combination with epirubicin. Therefore, we subsequently investigated its role and the underlying molecular mechanisms in respect to epirubicin-mediated anti-osteosarcoma. Our study indicated that overexpression of miR-1301 hinders cell proliferation and promotes apoptosis in osteosarcoma cells, which is similar to the functions of epirubicin. To gain a deeper insight into the molecular mechanisms of epirubicin and miR-1301 affecting apoptosis, the main regulatory molecules that govern the main basic mechanisms, such as the expression of apoptosis-related protein Bcl-2, Bax, cleaved-caspase-3, and poly (ADP-ribose) polymerase-1 (PARP1), were further examined. Bcl-2 is an anti-apoptosis protein, and dysregulation of Bcl-2-mediated apoptosis underlies a plethora of diseases.25 Bax is a pro-apoptotic member of the BCL-2 family.26 Also, detection of cleaved-caspase-3 in cells is an important method for apoptosis induced by a wide variety of apoptotic signals.2728 PARP1 is also a pro-apoptosis gene and could restore miR-520-induced cell apoptosis.29 Our findings demonstrated that epirubicin and upregulation of miR-1301 both reduces the expression of Bcl-2 and increases the expressions of Bax, cleavedcaspase-3, and PARP1, further reflecting the miR-1301-mediated pro-apoptosis effect. Importantly, we also revealed that miR-1301 enhances the function of epirubicin-induced anti-proliferation and pro-apoptosis in U2OS and SAOS-2 cells, indicating that miR-1301 increases the epirubicin effect. In contrast, miR-1301 knockdown attenuated the effect of epirubicin on osteosarcoma cells. Therefore, we inferred that miR-1301 facilitates epirubicin-mediated chemosensitivity in osteosarcoma.
miRNAs can regulate post-transcriptional gene expression and silence a broad set of targets genes. TRIAP1, a novel apoptosis inhibitor, has been shown to be upregulated in many types of cancers.30 Stable silencing of TRIAP1 induced late apoptosis in RPMI8226 cells.31 In addition, miR-18a inhibited ovarian cancer proliferation and induced apoptosis by targeting TRIAP1.32 miR-320b suppressed nasopharyngeal carcinoma cell proliferation and enhanced mitochondrial fragmentation and apoptosis by regulating TRIAP1. These data implied that TRIAP1 could be modulated by miRNAs. Here, we showed that miR-1301 repressed TRIAP1 expression by directly binding to the 3′UTR of TRIAP1. Moreover, the level of TRIAP1 was repressed by epirubicin in osteosarcoma cells, which was also rescued by introduction of miR-1301 inhibitors or TRIAP1 overexpression plasmid. Interestingly, restoration of TRIAP1 abolished the epirubicin-mediated cell proliferation inhibition. It is tempting to speculate that miR-1301 may participate in epirubicin-mediated chemosensitivity by targeting TRIAP1. Nevertheless, our study only investigated the role of miR-1301/TRIAP1 axis in vitro, and it needs to be further confirmed in xenograft models. Moreover, the role of TRIAP1 alone in osteosarcoma should be addressed in the future.
Taken together, our data highlighted the pivotal role of miR-1301 in epirubicin-mediated chemosensitivity in osteosarcoma. A novel miR-1301/TRIAP1 axis was first observed in osteosarcoma. Importantly, we found that an epirubicin-mediated miR-1301/TRIAP1 axis is involved in cell proliferation and apoptosis in osteosarcoma. Our findings provide powerful evidence of the molecular mechanism of chemosensitivity to epirubicin in osteosarcoma, which will help support clinical applications of a combination of epirubicin and miR-1301 in osteosarcoma.



 XML Download
XML Download