

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Canine influenza, a dog flu, is caused by two different influenza A viruses; equine-originated canine influenza virus (CIV) H3N8 and the avian-originated CIV H3N2 virus [123]. To date, it is yet to be reported on the transmission of CIV to human and CIV pandemic. However, recent findings on potential zoonotic transmission of CIV H3N2, genetic reassortment and fact of the quick and constant mutation in the influenza virus genome, CIV H3N2 may possibly lead to human threats in near future. This scenario emphasizes the need for rapid and effective CIV vaccine development to respond to the possible epidemic in the future [456].
At present, inactivated vaccines are produced either in egg or other animal cell systems, however, these conventional methods have shortcomings such as high production cost, long production time, and chance of human pathogen contamination, to name a few. Alternatively, the recombinant subunit vaccines from the plant-based production system may offer a novel platform for its safe, rapid, stable, consistence and scalable production. The vaccine production time could be further reduced by using instantly available plant biomass like lettuce leaves as a host cells. Also, lettuce can be used as an edible vaccine platform that does not require the need for trained personnel for immunization and cold chain distribution. The advancement of the transgenic lettuce produced hepatitis B surface antigen vaccine into a clinical trial (phase I) has proved the lettuce as an attractive and efficient host plant [7]. To date, several plant-made vaccines have been advanced to the different phases of a clinical trial. For example, Nicotiana benthamiana-produced human influenza virus-like particles (VLP) quadrivalent (phase III), human influenza H1N1 (phase I), tobacco produced Ebola vaccine (phase II), and norovirus capsid protein antigen in potato (phase I) [891011].
Influenza hemagglutinin (HA), a surface glycoprotein is the main target for subunit vaccine development because HA is the major viral antigen and it plays an important role in receptor binding and membrane fusion [12]. In addition, HA glycoprotein is also capable of assembling in VLP in plant by mimicking the native virus-like structure and it exhibits effective immunogenicity [1314]. HA vaccine with varying glycosylation pattern could be produced in plant by genetic engineering of the genes involved in glycosylation pathway. Many vaccine antigens for the various strain and subtype of human (H1N1, H3N2) and avian influenza (H5N1) have been expressed transiently in N. benthamiana and advanced to the different phases of clinical trial [15161718]. However, there is no report on the canine influenza vaccine antigen expressed in plant.
In this study, we investigated whether HA from Korean canine influenza strain A/canine/Korea/S3001/2015H3N2 can be expressed in N. benthamiana and edible leafy plant Lactuca sativa by using DNA viral vector systems. Recombinant HA (rHA) was successfully expressed in both the two plants. The fusion of endoplasmic reticulum (ER) retention signal, KDEL at C-terminal of HA sequence produced the basal-glycosylated rHA while the native signal peptide of HA at N terminus of HA produced the complex-glycosylated rHA in tobacco and lettuce.
Materials and Methods
Codon optimization and cloning
The amino acid and nucleotide sequence information were retrieved from the National Center for Biotechnology Information (NCBI), and Uniprot databases. The codons were first optimized to improve the expression in tobacco and lettuce. Since the codon usage by the specific amino acid in the human and within the different plant also vary, the codons were optimized based on the codon usage table of the N. benthamiana and L. sativa available at Kazusa DNA Research Institute (http://www.kazusa.or.jp). Moreover, the codon usage of highly expressed protein in plants, Rubisco and ubiquitin were analyzed in tobacco and lettuce using CAI calculator (http://genomes.urv.es/CAIcal/) and the codon of HA were adjusted. Importantly, the distribution of codon in the protein was maintained and the codon with zero usage frequency in the Rubisco and ubiquitin was avoided or used at a minimum. The optimized sequences of the full length of HA including the native signal peptide were synthesized (Macrogen, Seoul, Korea; http://www.macrogen.com). The synthesized HA DNA sequences were polymerase chain reaction (PCR) amplified by using the primer set HA3_SP_XhoI_F 5′-ATATCTCGAGATGAAGACTGTGATCGCCCTTTCTTATATTTTC-3′ and HA3_XbaI_R 5′-TTGTTCTAGAAATGCAAATGTTACATCTGATGTTGCCTC-3′ to generate HA with signal peptide and HA3_XhoI_F 5′-ATATCTCGAGATGCAGAACCTTCCAGGAAGTGAGAATAACGC-3′ and HA3_KDEL_XbaI_R 5′-TTGGTCTAGATCACAATTCATCCTTAATGCAAATGTTACATCTGATGTTGC-3′ to generated HA with KDEL at C terminus.
The amplified PCR product was digested with restriction enzymes XhoI and XbaI and ligated using the NEB T4 ligase (http://www.neb.com) into XhoI and XbaI site in geminiviral vector pBYR11ek2Md. For green fluorescent protein (GFP) control vector pBYR2ek2Md was used. The correct insertion of HA DNA was confirmed by colony PCR, restriction enzyme digestion, and Sanger sequencing.
Agroinfiltration procedure
Agrobacterium tumefaciens harboring the respective vector was cultured in yeast extract peptone medium containing kanamycin (50 mg/L), rifampicin (50 mg/L), and gentamicin (50 mg/mL) at 28℃ for overnight with 200 rpm to get the cell concentration around 1.6–1.8 optical density (O.D.). Bacterial cell density was determined by measuring O.D. at 600 nm. The agrobacterium cells were then collected by centrifugation at 5,000 rpm for 7 minutes at 4℃. Then, the pellet was re-suspended in the infiltration buffer (10 mM MES-KOH [pH 5.7], 10 mM MgSO4·7H2O) to give the O.D.600 of 0.2. N. benthamiana of 6- to 7-week-old, grocery-purchased red lettuce, and hydroponically-grown green lettuce were used for agroinfiltration. Agroinfiltration was done either by 1 mL syringe without needle for an individual leaf, or vacuum for the whole plant completely submerging them within a desiccator and applying vacuum for 2 minutes for tobacco and 5 minutes for lettuce at 70 mbar to allow the infiltration of agrobacteria into the apoplast spaces of submerged plant tissue. The leaf surface was washed after infiltration, then grown those infiltrated plants in 25℃, 50% humidity environment with a 16:00/8:00-hour day/night cycle.
Sodium dodecyl sulfate polyacrylamide gel electrophoresis and western blotting
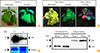
The leaves were harvested 4-day-after infiltration for protein detection and quickly frozen into liquid nitrogen and stored in −80℃ until further analysis. Hand-held ultraviolet lamp at 365 nm was used for visual detection of GFP in the plant (Fig. 1A, B). To obtain the expression levels of rHA, sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and western blot were carried out. For total protein extraction, agroinfiltrated leaves of 50 mg (fresh weight) were ground to the fine powder in liquid nitrogen and then mixed with 150–200 µL Laemmli loading buffer (50 mM Tris-HCl [pH 6.8], 2% sodium dodecyl sulfate, 5% beta-mercaptoethanol, 10% glycerol, 0.2 mg/mL bromophenol blue, 1× protease inhibitor cocktail [GenDEPOT, Katy, TX, USA]) followed by 10-minute incubation in boiling water and centrifuged for 10 minutes. Alternatively, 200–300 µL of extraction buffer (50 mM Tris pH 8.0, 150 mM NaCl, 2 mM ethylenediaminetetraacetic acid, 0.1% Triton X-100, 1× protease inhibitor cocktail [GenDEPOT]) was added to 100 mg and centrifuged at 13,000 rpm at 4℃ for 30 minutes. The supernatant was transferred to a new tube and protein concentration was determined by the Bradford assay. Total protein of 20 µg was resolved in the 10% Tris-glycine SDS-PAGE gel. The gel was then either stained with Coomassie brilliant blue G-250 or electrotransferred to polyvinylidene difluoride membrane (Merck, Darmstadt, Germany) using FLASBlot transfer buffer (https://advansta.com). The membrane was probed with a rabbit polyclonal influenza A H3N2-HA antibody (PA5-34930, Thermo Fisher, Waltham, MA, USA) in a dilution factor of 1:3,000 followed by the secondary antibody-alkaline phosphatase goat anti-rabbit IgG (AP-100-1, Vector Laboratories, Burlingame, CA, USA) in a dilution of 1:4,000 in phosphate buffered saline. Protein in the membrane was detected by using the ECL detection reagent. For the yield quantification, 100 ng and 125 ng commercially available H3N2-HA (recombinant partial length, Ab124562, Abcam, Cambridge, MA, USA) were used as a positive control. The plant made rHA yield was calculated by analyzing the protein band intensity in the membrane using the Image J software (National Institutes of Health, Bethesda, MD, USA) and compared with that of commercially available recombinant.
Results
Expression of canine HA (H3N2) antigen in Nicotiana benthamiana
The efficient production of a particular recombinant protein in a plant is a challenging because there are many combinatorial parameters governing the successful expression of the proteins. In order to find a right expression vector and the appropriate host plant that express the HA (H3N2) efficiently, we rationally chose DNA viral vector system and tested transient expression in the two host plants. In addition, to prevent the species-specific bias in the codon usage, synonymous mutations were introduced for optimal expression. So prior to cloning, we optimized the viral specific codon of HA from influenza A/canine/Korea/S3001/2015 (H3N2) towards the usage of preferred codon in tobacco and lettuce avoiding the use of rare codons that could negatively impact on HA expression.
To test whether the codon-optimized sequences of HA (H3N2) can be translated into recombinant protein in plant system, we expressed the HA gene in N. benthamiana, a well-established host plant for the production of the plant-based pharmaceutics. To this end, we cloned the full length of HA into the geminiviral vector expression system. The geminiviral vector expression system, pBYR11ek2Md is based on bean yellow dwarf virus (BeYDV), a DNA virus replicon system, to replicate huge amount of DNA template which produces the higher level of heterologous protein after transcription and translation in plant cell [192021]. The Agrobacterium tumefaciens harboring HA-carrying geminiviral vector was infiltrated into the tobacco leaves with the needless syringe or vacuum pump and the expression pattern was evaluated at the protein level. GFP-containing geminiviral vector, pBYR2eK2Md was also infiltrated as an expression control for the agroinfiltration procedure and monitor plant growth after infiltration (Fig. 1A). The extracted protein from the infiltrated leaves was resolved in SDS-PAGE gel and HA protein was detected by western blot analysis using rabbit polyclonal anti-influenza H3N2 HA antibody. The rHA was successfully expressed in tobacco leaves as revealed by the presence of strong protein band in the western blot (Fig. 1B).
Expression of canine HA (H3N2) antigen in Lactuca sativa
We then expressed rHA in an edible host plant lettuce which is amenable for the development of plant-produced oral vaccines. Lettuce offers an economical, quick and cold-chain free oral vaccine production platform. Although, transgenic expression system has been used to express the vaccine candidate protein against the human infectious viral disease in the lettuce, there is only a few report on transient expression of influenza vaccine candidate in lettuce [22232425]. Geminiviral vector system has also been applied to express the antibodies and antigens in transgenic system [202126].
We intended to express the rHA in lettuce because it can be further developed into a quick and inexpensive vaccine production system for pandemic and seasonal CIV. The production of vaccine antigen in lettuce could be especially more accountable at the time of emergency if we also could use the grocery lettuce for the transient production of a vaccine protein. Hence, we first tested whether geminiviral expression systems do express a recombinant protein in the lettuce bought from grocery stores. We vacuum-infiltrated the agrobacterium harboring the GFP in geminiviral vector (pBYR2eK2Md) into red lettuce head purchased from grocery store and green lettuce grown in the lab hydroponics system. The geminiviral vector system successfully worked for the GFP in the red lettuce head and green lettuce (Fig. 1C).
We further cloned the full length of the HA with the native signal peptide in pBYR11ek2Md vector and transiently expressed them in grocery-purchased lettuce (Fig. 1B). The expression level of rHA was determined 4-day-after infiltration by the western blot and probed with H3N2 HA-specific antibody. The rHA was successfully expressed in lettuce which was shown by the protein band in lettuce as similar to tobacco protein in the western blot; however, the higher level of rHA protein was produced in tobacco (Fig. 1B). We quantified the yield of rHA protein to be 9.3-13 mg/kg fresh weight of tobacco leaves (Fig. 1D).
Production of complex-glycosylated and basal-glycosylated rHA
Several studies suggest the importance of considering the glycosylation aspect for influenza vaccine design. During 2016–2017 influenza season, human H3N2 vaccine used at that time was not as effective as expected because of the loss of the HA glycosylation site at antigenic site B in the egg-produced vaccine, which emphasizes the need of considering protein glycosylation during vaccine design [2728].
In view of the possible impact of different glycosylation patterns of the rHA in its surface affinity, antigenicity, and efficacy of the vaccine, we were interested in producing CIV rHA with different glycosylation patterns. We fused the full length of rHA with the ER retention signal, KDEL to prevent the entry of rHA into the Golgi apparatus where the complex glycosylation occurs. To confirm and to compare the production of rHA without complex glycan by KDEL, we expressed the full length of rHA with native signal peptide as well (N-SP) (Fig. 2A). Geminiviral vector harboring rHA with native signal peptide or KDEL signal (KDEL) were infiltrated to both tobacco and lettuce leaves and the expressed proteins were analyzed in western blot. rHA protein was produced in the presence of either native signal peptide or KDEL signal in both tobacco and lettuce (Fig. 2B, C). However, the size of the rHA protein expressed with KDEL is smaller than that produced with the native signal peptide (Fig. 2B, C). KDEL signal retains rHA in the ER avoiding the further addition of the carbohydrate on rHA. In this case, the rHA protein size is proportional to the basal glycans being added in the ER. On the other hand, the native signal peptide with HA produced rHA protein of size around 75 kDa which is approximately 11 kDa bigger than the calculated size (64 kDa) of HA most possibly because of extra molecular weight of complex glycans added to HA by glycosylation in Golgi (Fig. 2B, C). We checked for the glycosylation sites in HA by using online glycosylation prediction site www.dtu.dk and found 6 N-glycosylation sites in asparagine residues (Fig. 2D) [29]. The similar nature of HA was reported for H5N1 where the observed protein size was bigger than the calculated when expressed full sequence in cytosol and apoplast [30]. It is interesting to observe the higher level of basal-glycosylated rHA production in lettuce than in tobacco, whereas higher level of complex-glycosylated rHA in tobacco (Figs. 1B, 2B, C).
Discussion
The goal of this study is to accomplish the transient expression of influenza HA antigen from A/canine/Korea/S3001/2015/H3N2 strain in the plant that can be developed into a vaccine. Multiple factors are responsible for the optimum expression of heterologous protein in the plant. We considered the species-specific codon usage, expression vectors system, secretion signal peptide, protein cellular compartmentation and host plant in our experiments. Viral codon of HA (H3N2) was modified to adjust codon usage by tobacco and lettuce on the basis of the codon usage table. The codon sequences not used by highly expressing protein Rubisco and ubiquitin were either avoided or maintained at a minimum in HA.
We investigated the expression system and host plant species to achieve the condition for the efficient expression of recombinant HA (H3N2) antigen in a plant. Geminiviral vector system successfully expressed the codon-optimized HA (H3N2) in N. benthamiana and L. sativa in our system (Fig. 1). The geminiviral vector system was found to be capable of inducing antibodies assembly and VLP formation in lettuce in addition to other different heterologous protein expression in tobacco [2021253132].
Our result showed that commercially available lettuce head can be used to produce influenza antigen when geminiviral vector system was employed (Figs. 1, 2). This platform carries a great value for the quick response to the influenza outbreak and pandemic owing to the rapid production of influenza vaccine reducing the time required for the preparation of host plant. Though tobacco performs better and is widely used for the production of plant-based pharmaceutical, the high level of nicotine and other secondary metabolite makes the downstream process of purification and regulatory approval necessary. So, we explored the use of the rapidly growing lettuce for geminiviral mediated transient expression which can be applicable for robust and safe vaccine production since lettuce is edible and generally regarded as safe, making the downstream purification simple and inexpensive. Lettuce has the potential for the development of needleless vaccine and cold-chainless vaccine distribution system. In support of this notion, lettuce produced exendin-4 (EX4) fused with cholera toxin B subunit has showed decreased glucose level in mouse when fed with a lyophilized leaf material [33]. Moreover, the lyophilized leaf material was not only rich in antigen concentration by 24-fold than in fresh leave but also stable up to 15 months at room temperature [33].
Our results showed that geminiviral vector system successfully expressed GFP and CIV rHA antigen in N. benthamiana and lettuce. The expression system of geminiviral vector and lettuce for the vaccine like CIV rHA could be easily advanced into large scale production system as the lettuce leaves are readily available through commercial grocery stores.
While employing a geminiviral vector expression system, native signal peptide at N terminus of HA resulted in higher level of rHA in lettuce and tobacco (Figs. 1, 2). Minimally-glycosylated rHA can be achieved by expressing the HA fused with ER retention signal KDEL which may possibly have different antigenicity than the original complex-glycosylated form (Fig. 2B, C). The rHA retained in ER presented the smaller size 64 kDa than 75 kDa. The size difference of rHA protein in two different cellular compartment may be accredited by the absence and presence of complex glycan. The glycosylation pattern analysis may provide direct evidence for the difference between the complex-glycosylated and basal-glycosylated proteins. To support our data, it was shown that the bigger HA protein size than calculated was observed when produced in the apoplast and smaller protein size when expressed in the chloroplast [30].
Influenza surface protein HA from different strains have a broad spectrum of glycosylation pattern that influences the antigenicity. In this perspective, plant-based vaccine production would be the relevant system to cope with seasonal and pandemic influenza since influenza HA vaccine with variable glycosylation pattern could be produced after the expression of HA antigen in the host plant with different degree of mutations in glycosylation-related genes. The vaccine with a broad spectrum of antigenicity could be produced transiently in N. benthamiana within the timeline of 3–4 weeks after the identification of the viral strain which may embrace a great value for the prompt response to novel influenza strain outbreak [3435].
In conclusion, we successfully produced complex-glycosylated and basal-glycosylated rHA antigen from A/canine/Korea/S3001/2015/H3N2 strain in N. benthamiana and L. sativa using geminiviral vector. Also, we provided the evidence that commercial lettuce could be employed as a host plant for the rapid production of vaccine that may help alleviate the situation at the time of influenza epidemic.

 XML Download
XML Download