

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Human periodontitis, also generally called periodontal disease, is a multifactorial polymicrobial infection characterized by a destructive inflammatory process that affects tooth supporting tissues and results in periodontal pocket formation, alveolar bone resorption, and consequently tooth loss [1]. During the onset and progression of periodontitis, the microbial community structure of the sub-gingival plaque changes, with an increase of proteolytic Gram-negative anaerobes including the black-pigmented rods in the genera Porphyromonas and Prevotella [2]. In particular, Porphyromonas gingivalis, Prevotella intermedia, and Prevotella nigrescens are known to be periodontopathic in humans [34].
As in humans, periodontitis is one of the most common disorders in dogs and cats. Approximately 80% of dogs and cats are estimated to experience some degree of periodontal disease until they are 4 years old [5]. The progression and clinical symptoms of periodontitis are almost identical in companion animals and humans. Various medical problems, such as anorexia, weight loss, chronic pain, sore or loose teeth, swollen gums, and loss of teeth, can occur in animals with periodontitis [67]. Furthermore, if left untreated, maxillary or mandibular bone can be destroyed, and bacteria from the periodontal pocket can enter the bloodstream, leading to renal, coronary, or hepatic diseases [8910]. The etiology of animal periodontitis is not fully described, but black-pigmented anaerobic bacteria have been isolated from the periodontal pockets of dogs and cats. Many of these isolates were Porphyromonas species including Porphyromonas gulae, Porphyromonas cangingivalis, and Porphyromonas cansulci [1112]. It has been reported that the proportion of Porphyromonas species isolated from canine plaque is about 6-fold higher at age 5 than at age 2 and correlates with disease severity [1314].
Inorganic polyphosphate (polyP) is composed of three to several hundred phosphate (Pi) residues linked by phosphohydride bonds [15]. The type of polyP varies depending on the length of the phosphate chain constituting it, for example, polyP3, polyP5, polyP15, polyP45, and so on. PolyP has long been used in the preparation of meat and dairy products due to its ability to improve binding properties and disperse proteins [16]. Some polyP, including sodium tripolyphosphate (Na5P3O10; polyP3), have been approved as food additives in the European Union (EU) and registered as Generally Recognized as Safe (GRAS) food additives by the FDA [171819]. The safety of polyP has been evaluated in various animal experiments. For example, polyP3 has been reported to be non-toxic when fed to dogs at 0.1 g/kg/day for 1 month [17]. PolyP is generally added to meat products at a concentration of 0.2%–0.3%, which is about half the upper limit defined by the EU (https://nofima.no/en/nyhet/2010/11/about-phosphate-and-meat/). PolyP also has the effect of preventing food spoilage because of its antimicrobial action against various Gram-positive bacteria and fungi [2021]. Interestingly, our recent studies have shown that polyP exerts excellent antimicrobial effects against black-pigmented Gram-negative anaerobes associated with human periodontal diseases such as P. gingivalis and P. intermedia [2223].
Periodontal disease can be prevented or effectively treated by inhibiting pathogenic microorganisms in the sub-gingival and supra-gingival plaques [724]. Because of its safety in use at the GRAS-level and its antimicrobial effect, we have focused on polyP3 as a potential therapeutic and preventive agent for periodontal disease in humans as well as in companion animals. Herein, we report for the first time the antibacterial effect of polyP3 against three Porphyromonas species implicated in periodontal disease in companion animals.
MATERIALS AND METHODS
Bacterial strains and culture condition
P. gulae KCTC5788, P. cansulci KCTC15047, and P. cangingivalis KCTC15089 were obtained from the Korean Collection for Type Cultures (Korea Research Institute of Bioscience and Biotechnology, Korea). The bacteria were grown on either Brucella agar (Becton, Dickinson and Company, USA) containing 5% laked sheep blood, 5 mg/L hemin, and 1 mg/L vitamin K1 (B-HK agar) or brucella broth containing 5 mg/L hemin and 1 mg/L vitamin K1 (B-HK) at 37°C anaerobically (80% N2, 10% H2, 10% CO2).
Determination of minimum inhibitory concentration (MIC)
PolyP3, purchased from Sigma Chemical Co. (USA), was dissolved in distilled water at 10% (wt/vol), filter-sterilized, and stored at −20°C until use. The MIC of polyP3 was determined by using the agar dilution method. Briefly, each bacterial culture growing exponentially was diluted to approximately 108 colony forming units per mL (CFU/mL) in fresh B-HK, as confirmed by viable bacteria counts. The agar plates containing polyP3 at 250–1,000 mg/L (corresponding to 0.025%–0.1%) were inoculated at approximately 105 CFU/spot and anaerobically incubated at 37°C for 3 days. MIC was defined as the lowest concentration that inhibited bacterial growth on the agar, in accord with CLSI guidelines [25].
Assessment of the viability of planktonic bacteria by CFU counting
Each bacterial culture suspension (500 μL/well), adjusted to approximately 2 × 105–2 × 106 CFU/mL in B-HK, was dispensed into a polystyrene 24-well plate containing B-HK (500 μL/well) supplemented with 0–750 mg/L polyP3. While the culture plates were incubated anaerobically at 37°C, 10 μL aliquots were removed from each well at 4-h intervals and inoculated onto agar plates, then incubated for 3 days.
Measurement of biofilm formation and biofilm viability
Bacterial culture suspension, adjusted to an optical density of 0.1 at 600 nm (OD600 = 0.1) in B-HK, was dispensed into a polystyrene 96-well plate (100 μL/well) containing B-HK (100 μL/well) with or without polyP3. As dead bacterial cells could attach to the surface of the polystyrene culture plates [2326], heat-killed (100°C for 10 min) bacterial cells were included as controls. After 24 h of incubation, planktonic and loosely bound bacterial cells were removed by aspiration of the spent media. The biofilms remaining on the surface were washed twice with saline and stained with 0.1% crystal violet (CV) for 10 min. After washing three times with distilled water and air drying, the bound dye was solubilized in 95% ethanol (200 µL/well), diluted 4-fold, and the OD600 recorded. The biofilm viability was measured using a LIVE/DEAD BacLight Bacterial Viability Kit (Invitrogen, USA) according to the manufacturer's instructions.
Scanning electron microscopy (SEM)
The bacterial biofilms were developed in the wells of 24-well polystyrene plates for 24 h with or without polyP3, as described above. Biofilm specimens for SEM were prepared as described previously [26]. Following critical point drying, the specimens were coated with gold, then observed under a scanning electron microscope (Model S-4700; Hitachi High Technologies America, Inc., USA).
Assessment of nucleotide leakage
The bacterial culture suspension was adjusted to an OD600 = 0.1 in B-HK and further incubated at 37°C for 24 h with or without polyP3, then centrifuged. The nucleotides contained in 1 mL of the supernatant were concentrated by ethanol precipitation, then dissolved in 50 µL of distilled water. Nucleotides in the solution were electrophoresed on an agarose gel and visualized under ultraviolet light after ethidium bromide (EtBr) staining.
RNA extraction and sequencing
P. gulae was chosen for further study. Exponentially growing P. gulae cells were transferred to fresh B-HK and incubated for 80 min with or without 500 mg/L polyP3, then harvested. RNA was extracted using TRIzolTM (Invitrogen, USA) and cleaned with RNeasy® Plus Mini Kit (QIAGEN, Germany). Ribosomal RNA was removed by using a Ribo-Zero rRNA Removal Kit for Bacteria (Illumina, USA). A standard sequencing system protocol (TruSeq Stranded Total RNA prep; Illumina) was used to make the cDNA libraries for RNA sequencing (RNA-Seq). The cDNA fragments (around 300 bp) were isolated using gel electrophoresis, amplified by PCR, and sequenced on an Illumina HiSeq 2000 in the paired-end sequencing mode (2 × 101 bp reads). The above procedures were repeated for two independent biological replicates of P. gulae.
RNA-Seq raw read processing and analysis of differential gene expression
Quality of the raw sequencing reads was assessed, and the qualified raw sequencing reads were aligned to the P. gulae DSM 15663 draft genome (NCBI RefSeq assembly accession: GCF_000378065.1) using the Bowtie2 alignment tool (version 2.2.9). Uniquely and properly mapped read pairs were used for further analysis. Gene annotation information was downloaded from NCBI RefSeqASM37806v1 (https://www.ncbi.nlm.nih.gov/assembly/GCF_000378065.1/). The reads per kilobase million (RPKM) measurement unit was used to evaluate gene expression levels. To identify differentially expressed genes (DEGs) between polyP3 exposed and unexposed bacterial samples, the DESeq2 R package was used. Differential expression was defined as at least a 4-fold change between a pair of samples with a false discovery rate of 1% for genes expressed at ≥3 RPKM in ≥1 sample(s). RNA-sequencing data were submitted to the Gene Expression Omnibus with ID GSE93901.
Functional annotations and functional enrichment analysis
The cluster of orthologous (COG) information was downloaded from the NCBI COG database. To further extend the comprehensive functional annotation and orthologous gene information, 1,870 NCBI RefSeq annotated gene FASTA sequences were submitted to the eggNOG-mapper web service (http://eggnogdb.embl.de/app/emapper#/app/emapper). Using clusters and phylogenies from the eggNOG database, functional annotations of 1,647 genes were generated based on orthology assignments and used in the gene ontology (GO) term and KEGG pathway enrichment test, as described previously [27].
RESULTS AND DISCUSSION
PolyP3 killed Porphyromonas species
By applying an agar dilution method, MICs of polyP3 against P. gulae, P. cansulci, and P. cangingivalis were determined to be 750, 750, and 500 mg/L, respectively (Fig. 1A). We further monitored the antibacterial effect of polyP3 on planktonic bacterial cells in a liquid medium by counting viable cells at 4-h intervals for 24 h. The CFUs of P. gulae exposed to ≥ 250 mg/mL polyP3 and P. cansulci exposed to ≥ 500 mg/L polyP3 decreased to less than 0.1% of the initial inoculums within 24 h (Fig. 1B and 1C). P. cangingivalis was the most susceptible to polyP3 among the three species and was completely killed by 250 mg/L polyP3 within 24 h (Fig. 1D). That is, polyP3 exerted bactericidal activity (defined as a ≥ 99.9% killing rate within 24 h [25]) against planktonic cells of all three Porphyromonas species at lower concentrations than the MICs determined by the agar dilution method. Similarly, the MIC of polyP against the human periodontopathic P. gingivalis and P. intermedia was 600–750 mg/L by the agar dilution method, whereas polyP in the liquid medium showed a bactericidal effect against the two bacteria at sub-MIC levels [2223]. Some antimicrobials, including polyP, may exert a higher effect in liquid media than in solid media, presumably because the liquid medium provides an environment favorable for the migration and diffusion of the antimicrobial agents, allowing easy contact with individual bacterial cells.
Fig. 1
Antibacterial effect of polyP3 against P. gulae, P. cansulci, and P. cangingivalis. (A) MIC determination of polyP3 obtained by agar dilution method. The bacterial cells were spot-inoculated (approximately 105 CFU/spot) onto brucella blood agar plates containing polyP3 at various concentrations and incubated at 37°C for 3 days anaerobically. The MIC was defined as the lowest concentration that inhibited the bacterial growth on the plate. (B) Time-kill curve of polyP3 against the Porphyromonas species in the liquid medium. Results are presented as means ± standard deviations from three independent experiments.
P. gulae, Porphyromonas gulae; P. cansulci, Porphyromonas cansulci; P. cangingivalis, Porphyromonas cangingivalis; MIC, minimum inhibitory concentrations; CFU, colony forming units.

PolyP3 reduced biofilm formation by Porphyromonas species
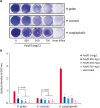
In the absence of polyP3, P. cangingivalis was observed to form the most abundant biofilm for 24 h, followed by that of P. gulae and P. cansulci (Fig. 2). It was also observed that some of the heat-killed P. gulae, P. cansulci, and P. cangingivalis adhered to the polystyrene surface. Biofilm formation was significantly reduced by polyP3 at concentrations that killed the planktonic bacteria. When exposed to polyP3 (250–750 mg/L), the biofilm formation by P. gulae decreased by 57.9%–78.5%. The biofilm formation by P. cansulci was reduced by 41.2%–51.6% when exposed to polyP3 (500–750 mg/L). P. cangingivalis, which has the highest biofilm-forming ability, showed the most remarkable reduction of the biofilm formation by 81.6%–86.0% in the presence of polyP3 (250–750 mg/L).
Fig. 2
Effect of polyP3 on biofilm formation by P. gulae, P. cansulci, and P. cangingivalis. The bacterial culture was diluted to an OD600 = 0.1 in B-HK and further incubated in 24-well plates with polyP3 at the indicated concentrations for 24 h. Heat-killed bacterial cells that were initially killed by exposure to 100°C for 10 min were included as controls. The biofilm biomass was quantitated by CV staining and expressed as OD600. (A) Images of the biofilms stained with CV. (B) Quantitation of the biofilm biomass. Data presented are means ± standard deviations from two independent experiments performed in triplicate.
*p < 0.05, significant difference observed when compared with the controls (in the absence of polyP3) in one-way analysis of variance with Tukey's post-hoc tests. All statistical analyses were performed using GraphPad Prism statistical software (version 5.01; GraphPad, USA).
P. gulae, Porphyromonas gulae; P. cansulci, Porphyromonas cansulci; P. cangingivalis, Porphyromonas cangingivalis; OD, optical density; CV, crystal violet.
PolyP3 caused various degrees of membrane disruption in Porphyromonas species
We observed the morphology of the biofilms and individual bacterial cells by SEM. After 24 h, the three Porphyromonas species formed multi-layered biofilms on the polystyrene surface of the culture plate (Fig. 3). As the concentration of polyP3 increased from 250 to 750 mg/L, the biofilm cell density of P. gulae and P. cansulci decreased remarkably. The biofilm cell density of P. cangingivalis also decreased notably, but a polyP3 concentration-dependent change was not apparent in the range of 250–750 mg/L. These results are consistent with the quantitative changes in the biofilm biomass measured by CV staining (Fig. 2). Morphological changes of individual cells in the biofilm following polyP3-exposure were not observed in P. gulae and P. cansulci. In contrast, the morphology of P. cangingivalis in the biofilm markedly changed in the presence of 250–750 mg/L polyP3, accompanied by distinct individual cell destruction. Notably, the biofilm viability of P. cangingivalis was reduced more than 2-fold when treated with 500–750 mg/L polyP3 than when treated with 250 mg/L polyP3 (Fig. 4A), unlike the above results (Figs. 2 and 3) where polyP3 concentration-dependent changes were not apparent.
Fig. 3
SEM images of P. gulae, P. cansulci, and P. cangingivalis. Each bacterial culture was diluted to an OD600 = 0.1 in B-HK broth and then further incubated in 24-well plates with polyP3 at the indicated concentrations for 24 h. Heat-killed bacterial cells that were initially killed by exposure to 100°C for 10 min were included as controls. The biofilms were observed by SEM at a magnification of 20,000× and 100,000× operating at 5 kV.
SEM, scanning electron microscopy; P. gulae, Porphyromonas gulae; P. cansulci, Porphyromonas cansulci; P. cangingivalis, Porphyromonas cangingivalis; OD, optical density.

Fig. 4
Biofilm viability and bacterial membrane leakage. The bacterial culture adjusted to an OD600 = 0.1 in B-HK was further incubated with polyP3 at the indicated concentrations for 24 h. (A) Viability of P. cangingivalis biofilms was measured using a LIVE/DEAD BacLight Bacterial Viability Kit. The percentage of biofilm viability was determined relative to the control.
*p < 0.02, significant difference between the experimental groups in one-way analysis of variance with Tukey's post-hoc tests. (B) Nucleotides leaking into the bacterial culture supernatant. The nucleotides contained in 1 mL of the culture supernatant were concentrated, electrophoresed on agarose gel, stained with EtBr, and visualized under ultraviolet light. Band intensity was analyzed with ImageJ version 1.49j (National Institutes of Health, USA).
P. gulae, Porphyromonas gulae; P. cansulci, Porphyromonas cansulci; P. cangingivalis, Porphyromonas cangingivalis; OD, optical density.

If a bacterial membrane was damaged by an antimicrobial agent, intracellular components such as small ions, proteins, and nucleotides would have leaked. Hence, quantitative changes in the nucleotides leaked from bacterial cells are a good indicator of the extent of membrane damage [28]. To further evaluate how much polyP3 damages the bacterial membranes of the three species, we used the supernatant from the bacterial culture to measure nucleotides released from the cytoplasm. EtBr-stained materials were scarce in the culture supernatant of P. cansulci regardless of polyP3 treatment (Fig. 4B). In contrast, EtBr-stained materials leaked from P. gulae increased by about 1.4- to 1.5-fold by 250–750 mg/L polyP3 treatment, indicating that polyP3 caused damage to some extent to the bacterial membrane of this species. As predicted by severe morphological changes, nucleotide leakage due to polyP3 exposure was most prominent in P. cangingivalis, showing 1.6- and 5.9-fold increases by 500 and 750 mg/L polyP3, respectively.
PolyP has been reported to chelate divalent cations present in Gram-positive bacterial membranes and cause direct cell lysis [21], but not to induce cell lysis of Gram-negative bacteria [2226]. Our previous studies demonstrated that polyP has a very strong bactericidal effect on Gram-negative bacteria, P. gingivalis and P. intermedia, without causing significant membrane damage or cell lysis [2226]. Herein, we report the first case of a direct-damaging effect of polyP against Gram-negative bacteria, demonstrating that polyP3 induces nucleotide leakage from P. cangingivalis as well as severe membrane alteration in that bacterium. Interestingly, the degree of membrane damage in the three Porphyromonas species caused by polyP3 varied. Considering that different bacterial species belonging to the same genus can have different envelope components [28], it is presumed that P. cangingivalis can be highly sensitive to polyP3 because the target components of polyP3, such as cations, have a particularly important role in cell membrane stability of this bacterium. Membrane destruction by polyP3 may have resulted in the release of a variety of cellular components, which may increase aggregation between dead and living cells of P. cangingivalis, causing an increase of the biofilm biomass. As a result, the biofilm cell density of P. gingivalis, as observed by SEM, was affected little by polyP3 even at higher concentrations. (Figs. 3 and 4). On the other hand, polyP3 showed a strong antimicrobial effect against P. gulae and P. cansulci with little or no cell membrane destruction, indicating that other antimicrobial mechanisms of polyP3 act on these bacteria rather than by a direct attack on the membrane.
PolyP3 caused extensive changes in the P. gulae transcriptome
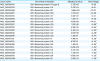
Of the Porphyromonas species found in companion animals, P. gulae has the highest sequence identity (up to 98%) with P. gingivalis [12], a "keystone pathogen" that affects human oral microbiota and has an important role in the development and progression of human periodontitis [29]. In addition, P. gulae is the second most abundant Porphyromonas following P. cangingivalis in dogs with periodontitis [30]. To better understand the mechanism of antimicrobial action of polyP3, P. gulae mRNA was sequenced. We obtained 50 million pairs of reads (2 × 101 bp) using the standard paired-end RNA-Seq protocol on the Illumina Hi-Seq 2000 sequencer. Table 1 shows the read counts and mapping results of the control and experimental groups. Approximately 82% (41 million) of the read pairs were uniquely mapped. The 1,983 genes of the draft genome of P. gulae DMS 15663 (NCBI RefSeq annotation, March 14, 2016) have at least one read mapped.
Table 1
RNA-Seq read mapping summary

Analysis of the DEGs showed that polyP3 causes significant biological changes in P. gulae. A total of 603 genes (about 30% of the P. gulae genome) were differentially expressed by polyP3 (> 4-fold, adjusted p value < 0.01) (Fig. 5 and Supplementary Table 1). In both the GO and the KEGG pathway analyses, the common DEGs with down-regulated expression trends were significantly enriched in thiamine biosynthesis, peptidoglycan metabolism, and ubiquinone biosynthesis (Table 2).
Fig. 5
Differential gene expression between control and polyP-treated groups. A volcano plot shows the DEGs. Colored dots indicate the significantly over-expressed genes in each sample: red, up-regulated genes; blue, down-regulated genes. The x-axis indicates the magnitude of fold changes (log2 transformed) and the y-axis indicates the adjusted p value (−log2) obtained by Benjamini-Hochberg correction.
FC, fold change (control/experimental).
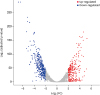
Table 2
Differentially expressed genes related to metabolism of thiamine, ubiquinone, and peptidoglycan

PolyP3 hindered energy metabolism and peptidoglycan biosynthesis of P. gulae
Thiamine, or vitamin B1, is vital for the survival of all living things. Thiamine pyrophosphate (TPP), a biologically functional form of thiamine, acts as a cofactor for several enzymes in key cellular metabolic pathways such as glycolysis, pentose phosphate pathway, and the citric acid cycle [31]. The de novo biosynthesis of thiamine is found in prokaryotes, fungi, and plants, but animals must obtain it from a dietary source [32]. Thus, the thiamine biosynthetic pathway is considered a potential antibiotic target [32]. Genes involved in thiamine biosynthesis are functionally regulated by the TPP riboswitch, a TPP-sensing RNA located in the untranslated region of mRNA [33]. Two regulatory mechanisms based on the structure of TPP riboswitches and their ligands are premature transcription termination and translation inhibition [34]. Using the Pfam database [35], we found that P. gulae has two TPP riboswitch motifs (Pfam ID: RF00005) and that seven thiamine biosynthesis-related genes are downstream of the TPP riboswitches. Among these genes, expressions of the six downstream genes, except the most upstream ThiS-encoding gene, were significantly reduced in P. gulae exposed to polyP3 (Table 2). These results suggest that premature transcription termination by a TPP riboswitch could be a crucial factor in the down-regulation of thiamine biosynthesis-related genes and that polyP3 may have affected this transcription termination process. Further research is needed to determine the exact mechanism by which polyP3 affects thiamine metabolism in P. gulae. Notably, polyP3 also significantly reduced the expressions of several P. gulae genes involved in the biosynthesis of quinone (Table 2), an essential lipophilic carrier in the electron transport chain for ATP production [2736]. Taken together, the results strongly suggest that the antimicrobial activity of polyP3 against P. gulae is closely related to an energy metabolism disorder through the inhibition of endogenous thiamine and quinone biosynthesis.
Energy metabolism disturbances by polyP3 are expected to be closely linked to interference with cell wall biosynthesis and protein synthesis. Indeed, the genes important for peptidoglycan synthesis were significantly down-regulated by polyP3 (Table 2). The decrease in peptidoglycan synthesis would have contributed to the cell wall instability of P. gulae, probably causing a nucleotide leak from the bacterial cytoplasm, as shown in Fig. 4. Very similar results were observed in our previous study on P. gingivalis; polyP75 significantly reduced the expression of a series of genes involved in the energy metabolism and cell envelope synthesis in P. gingivalis [27]. However, no apparent cell membrane lysis was observed in P. gingivalis exposed to polyP75 [22] as well as in P. cansulci exposed to polyP3 (Figs. 3 and 4). The differences in the degree of cell membrane damage by polyP may have been due, at least in part, to the type, concentration, and exposure time of polyP, as well as the different cell envelope composition among the species.
Another interesting point is the increased expression of many genes encoding ribosomal proteins in P. gulae exposed to polyP3 (Table 3), similar to that observed in P. gingivalis [27]. The production of ribosomes in fast-growing bacteria consumes about 40% of the total energy and is, therefore, strictly controlled at several steps [37]. When ribosomes are damaged because of poor growing conditions or exposure to antibiotics, bacteria can to some extent increase the synthesis of ribosomal proteins and accumulate ribosome precursors to compensate for the slowed function [383940]. Thus, it can be assumed that up-regulation of the genes encoding ribosomal proteins in P. gulae and P. gingivalis exposed to polyP may reflect a compensatory response to slower or disturbed function of the ribosome due to energy metabolism disorders.
Table 3
Differentially expressed genes related to ribosome
Collectively, polyP3 showed excellent in vitro antimicrobial activity against the Porphyromonas species tested in this study by disturbing the cell membranes and/or by inhibiting energy metabolism and cell envelope biosynthesis. Taken together, the results suggest that polyP3, with proven safety as a food additive, can be an attractive and promising candidate for use in the treatment and prevention of periodontal disease in companion animals. However, dental biofilm is more complex than the single bacterial species cultivated in laboratories, and most of the tissue destruction in periodontal disease is related to the host’s inflammatory response to microbial infection. Hence, antimicrobial activity of polyP3 against other periodontal pathogens of companion animals and its effect on the host inflammatory response need to be evaluated before applying polyP3 to the treatment and prevention of periodontal diseases.
 XML Download
XML Download