

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Atopic dermatitis (AD) is a common inflammatory cutaneous disorder1 caused by complex interactions among environmental factors, changes in skin barrier function, and immune system.23 AD is a risk factor that can predispose an individual to allergic diseases such as asthma or rhinitis45 since disruption of the skin barrier in AD patients can lead to allergen sensitization, which can, in turn, induce allergic esophageal inflammation or food anaphylaxis.67
A number of studies have suggested that an imbalance in T helper (Th) 1/Th2 cells and skin barrier dysfunction contribute to AD pathogenesis.8 Defective skin barrier causes unregulated activation of proteases in the epidermis, leading to the production of Th2-type immune-inducing factors. AD is dominantly mediated by Th2 cells910 and is associated with increased immunoglobulin (Ig) E and the production of Th2 cytokines, such as interleukin (IL)-4 and IL-13 in AD lesions.111213 Moreover, there is growing recognition that AD is not only associated with Th2 immune response, but that Th17 and Th22 inflammatory responses are also involved.141516 Activated T cells are recruited to the skin in acute and chronic cutaneous inflammatory diseases.17 CD4+ T cells are key factors in allergic inflammatory diseases. Several studies have shown that inhibition of CD4+ T-cell activation inhibited Th1, Th2 and Th17 cell differentiation with capsiate treatment.18 These immune responses require specific signals for T-cell activation that engage receptor systems on T cells and dendritic cells (DCs). The initial interaction involves contact between T-cell receptor and the antigenic peptide bound to major histocompatibility complex class II on the DC surface,19 followed by adhesion and expression of co-stimulatory molecules on T cells (CD28, cytotoxic T-lymphocyte-associated protein-4 and CD6) and DCs (CD80/B7.1 and CD86/B7.2).20
Activated leukocyte cell adhesion molecule (ALCAM) is a transmembrane protein of the Ig superfamily2122 that is expressed on a variety of cell types. CD6 is highly expressed on T cells and binds to ALCAM on DCs.23 Crosslinking of CD6 and ALCAM contributes to both early and later stages of T-cell proliferation,24 suggesting that the CD6-ALCAM interaction is necessary for maximum antigen-specific T-cell activation. Indeed, blocking this interaction with an antibody suppressed IL-2 production.25 Lymphocyte function-associated antigen 1 (LFA-1) and intercellular adhesion molecule (ICAM)-3 are also involved in effector T-cell function26; although this is similar to those of LFA-1 and ICAM-3 localized in the synapse, CD6-ALCAM accumulation at this site occurs independently of these adhesion molecules.23 Another study reported that the engagement of CD6 with ALCAM at the synapse provided the co-stimulatory signal for activation of γδ T cells in response to cell-associated non-peptide antigens in a way that was similar to activation of αβ T cells by peptide antigen-loaded DCs, which involved immunological synapse stabilization.27 It is thus likely that the CD6-ALCAM interaction affects diverse subsets of T cells.
Prior studies demonstrated that ALCAM contributes to ovalbumin (OVA)-induced allergic asthma and food allergy by stimulating T-cell activation and proliferation.2829 Although ALCAM is associated with hypersensitivity and T cell activation in the initiation of immune responses, its function in AD has not yet been demonstrated. Therefore, the present study examined the role of ALCAM in AD pathogenesis using an OVA-induced AD-like allergic skin inflammation mouse model and compared ALCAM-deficient and wild-type (WT) mice in terms of molecular and histological changes.
MATERIALS AND METHODS
Subjects
ALCAM levels were measured in the serum of children who visited the allergy clinic at Severance Children's Hospital for AD or for a general health checkup between June 2010 and November 2014. Children with AD fulfilled the revised Hanifin and Rajka criteria30 and were subdivided into the following 3 groups according to disease severity based on the SCORing AD (SCORAD) index:3132 mild (SCORAD index ≤ 25), moderate (25 < SCORAD index ≤ 50) and severe (SCORAD index > 50). They had no symptoms of other allergic diseases such as asthma or allergic rhinitis. Healthy controls had no history of AD or other allergic diseases such as asthma, allergic rhinitis, or inflammatory disease. At the first visit, blood samples were obtained from all subjects. Eosinophil count was determined using a hematological analyzer (NE-8000; Sysmex, Kobe, Japan), and IgE level was measured with the CAP assay (Pharmacia, Uppsala, Sweden). The study protocol was approved by the Institutional Review Board (IRB) of Severance Hospital, and written informed consent was obtained from subjects or their parents (IRB No. 4-2004-0036).
Skin biopsy samples were obtained from 1 healthy control and 1 AD patient who were diagnosed according to the criteria of Hanifin and Rajka.33 This study was approved by the IRB of Yonsei University Health System, Severance Hospital (No. 4-2013-0624), and informed consent was obtained from all subjects before they participated in the study. This study protocol was performed in accordance with the ethical guidelines of the Korean Bioethics and Safety Act.
Mice
C57BL/6 (B6) mice were purchased from Orient Bio (Sungnam, Korea). ALCAM-deficient mice on the B6 genetic background were purchased from Jackson Laboratory (Bar Harbor, ME, USA). Mice were maintained under pathogen-free conditions with controlled humidity and temperature. Animal experiments in this study were carried out in accordance with the Laboratory Animals Welfare Act, the Guide for the Care and Use of Laboratory Animals and the Guidelines and Policies for Rodent experiment provided by the Institutional Animal Care and Use Committee (IACUC) and were approved by the IACUC of the Yonsei University Health System (reference No.2014-0282).
Induction of AD
The OVA-induced AD model was established as previously described.34 Mice were anesthetized, and their back skin was shaved and tape-stripped 5 times with cellophane tapes (Nichiban, Tokyo, Japan). For epicutaneous (EC) sensitization, 100 μg OVA (grade V; Sigma-Aldrich, St. Louis, MO, USA) in 100 μL of normal saline was placed on a 1×1 cm2-patch of sterilized gauze attached to the dorsal skin with Tegaderm (1,624W; 3M Health Care, Maplewood, MN, USA). Mice were thus subjected to a total of three 1-week EC sensitizations with a 2-week interval between individual sensitization periods. Transepidermal water loss (TEWL; Delfin, Kuopio, Finland) and clinical score were assessed during the experimental period.
To induce oxazolone (OXA)-AD-like model, the dorsal skin of mice was sensitized with 50 μL of 3% OXA (Sigma-Aldrich) at day 0. After 5 days, the ear was challenged by 20 μL of 0.6% OXA every other day until day 13. TEWL, clinical score and ear thickness (Mitutoyo, Kanagawa, Japan) were measured during the experimental periods. On day 14, mice were sacrificed to collect the skin biopsy specimen and blood.
The clinical severity of skin lesions was scored according to the macroscopic diagnostic criteria that were used for the NC/Nga mouse. The severity of dermatitis was evaluated once a week, just before each elicitation. The development of 1) erythema/hemorrhage, 2) scarring/dryness, 3) edema and 4) excoriation/erosion was scored as 0 (none), 1 (mild), 2 (moderate) and 3 (severe). The average of the individual scores was taken as the clinical score.35
Quantitative real-time polymerase chain reaction (PCR)
AD skin lesion samples were obtained after the last sensitization to assess messenger RNA (mRNA) expression levels. Total RNA was isolated with TRIzol reagent (Invitrogen, Carlsbad, CA, USA), and 2 μg were used to synthesize complementary DNA using superscript II reverse transcriptase (Invitrogen). Quantitative real-time PCR was performed on an Applied biosystem (Foster City, CA, USA) and target gene levels were quantified with AccuPower Greenstar qPCR PreMix (Bioneer, Daejeon, Korea) according to the manufacturer's instructions. Forward and reverse primers for target gene amplification were synthesized by Integrated DNA Technologies (Coralville, IA, USA). The primer details are shown in Table 1. Target gene mRNA levels were quantified relative to that of β-actin.
Table 1
Sequences of the primers used for quantitative real-time polymerase chain reaction

![]()
Enzyme-linked immunosorbent assay (ELISA)
Mice were bled by cardiac puncture and serum was collected by centrifugation and stored at −70°C until use. Serum levels of total IgE were quantified with a specific ELISA kit (BD Biosciences, San Diego, CA, USA) according to the manufacturer's protocol. ALCAM levels in mouse skin homogenate were quantified by ELISA (R&D Systems, Minneapolis, MN, USA) according to manufacturer instructions.
Hematoxylin and eosin (H&E) staining
Dorsal skin was fixed in 10% buffered formalin, and samples were embedded in paraffin and cut into sections at a thickness of 4 μm that were mounted onto slides and stained with H&E. Histological changes were scored (0−4) according to the degree of inflammation, immune cell infiltration, edema and epithelial hyperplasia by visualization under a light microscope (BX40; Olympus, Center Valley, PA, USA).35
Electron microscopy
Skin samples were pre-fixed in Karnovsky's fixative solution (2% glutaraldehyde-paraformaldehyde in 0.1 M phosphate buffer, pH 7.4) for 7 days, then rinsed with phosphate-buffer saline and pre-fixed with 1% osmium tetroxide. Samples were dehydrated in a graded series of ethanol, then infiltrated with propylene oxide and embedded using the Ply/Bed 812 kit (Polysciences, Warrington, PA, USA). Thin sections (70 nm) were cut and stained with 7% uranyl acetate and lead citrate and observed under a JEM-1011 transmission electron microscope (JEOL, Tokyo, Japan). Images were obtained with a Mega View III camera (Olympus, Tokyo, Japan).
Flow cytometry
Cell suspensions were isolated from the skin and skin draining lymph nodes (LNs) of mice. To collect whole skin cells, skin was detached from the body and incubated for 1 hour in Roswell Park Memorial Institute (RPMI) 1640 medium (Hyclone, Logan, UT, USA) containing 1 mg/mL dispase II (Sigma-Aldrich). The samples were cut into small pieces and incubated for 1.5 hours in RPMI 1640 medium containing 10% fetal bovine serum (FBS), 0.8 mg/mL collagenase type II (Worthington Biochemical, Lakewood, NJ, USA), and 50 μg/mL DNase I (Roche, Freehold, NJ, USA).36 The LNs were cut into small pieces and digested in collagenase type II (0.5 mg/mL; Worthington Biochemical) or DNase I (Roche; 0.02 mg/mL) at 37°C for 45 minutes. Digested skin draining LNs and skin were passed through a 40-µm cell strainer (BD Biosciences). Red blood cells were lysed using ammonium chloride-potassium lysing buffer, and the remaining cells were washed and resuspended in phosphate-buffered saline (PBS) containing 5% FBS.
Single-cell suspensions were then stained with antibodies for flow cytometry. The following antibodies were used: CD3 (17A2), CD4 (GK1.5), CD44 (IM7), CD62L (MEL-14) and isotype controls (rat IgG) (eBioscience, San Diego, CA, USA). Fixable Viability Dye (eBioscience) was used to exclude dead cells. Stained cells were analyzed by flow cytometry using an LSR II (BD Biosciences), and data were analyzed with Flow Jo software (Tree Star, Ashland, OR, USA).
Statistical analysis
Continuous data were tested for normality using the Kolmogorov-Smirnov test or Shapiro-Wilk test and reported accordingly as the mean ± standard error of the mean. The Student's t-test and 1-way analysis of variance were used to evaluate differences between continuous variables. Categorical data are presented as counts and percentages, and the χ2 test was used for comparisons. Correlations between ALCAM level and SCORAD index were evaluated by Pearson correlation analysis. Statistical analyses were performed with SPSS v.20 software (SPSS Inc., Chicago, IL, USA). Differences were deemed significant at P < 0.05.
RESULTS
ALCAM levels are altered in serum from human AD and OVA-induced AD model
The clinical characteristics of the study subjects are summarized in Table 2. The AD group was significantly younger in age and included more males than the control group. Serum total eosinophil count and IgE level were higher in the AD group than in healthy controls, and the former also showed elevated serum ALCAM levels (26 ± 3.4 ng/mL vs. 30.8 ± 3.5 ng/mL, P < 0.001) (Fig. 1A) even after adjusting for age and sex (β = 3.68, P < 0.001). Moreover, ALCAM levels in children with AD tended to increase with AD severity (Fig. 1B), as evidenced by the positive correlation with the SCORAD score (r = 0.392, P < 0.001) (Fig. 1C).
Table 2
Characteristics of the study population

Values represent mean ± standard deviation or number (%).
*P < 0.05; †P < 0.001 vs. healthy control (Student's t-test).
![]()
 | Fig. 1ALCAM expression in pediatric AD patients and OVA-induced AD mice. (A-C) ALCAM level was measured by ELISA in serum samples from healthy controls (n = 44) and AD patients (n = 114). Pirate plots show the distribution of ALCAM levels for each group; horizontal lines represent means and boxes represent 95% confidence intervals. (A) ALCAM level in human serum. (B) Increase in serum ALCAM level as a function of AD severity increases. (C) Correlation between serum ALCAM level and AD SCORAD index. (D) Serum ALCAM abundance was assessed by ELISA.The data represent mean ± standard error of the mean.
ALCAM, activated leukocyte cell adhesion molecule; AD, atopic dermatitis; OVA, ovalbumin; PBS, phosphate-buffered saline; SCORAD, SCORing atopic dermatitis; ELISA, enzyme-linked immunosorbent assay.
*P < 0.05; †P < 0.001 by t-test (n = 5–10 mice/group).
|
To examine whether development of OVA-sensitized AD can regulate the levels of ALCAM in WT mice. OVA-sensitized mice showed increased serum ALCAM level (Fig. 1D) compared to PBS mice.
ALCAM deficiency alleviates skin barrier disturbance
One of the common features of AD is an impaired skin barrier. In this AD model, clinical score and TEWL level, which shows the abnormal skin barrier function, was significantly increased from day 7 in WT/OVA mice (Fig. 2A and B). However, ALCAM−/−/OVA mice had markedly decreased clinical score and TEWL compared to WT mice (Fig. 2A and B). We next examined lamellar body (LB) morphology and permeability barrier gene expression in ALCAM−/− and WT mice. LB is a unique organelle that delivers lipids to the stratum corneum. Failure of LB secretion was previously reported as a feature of AD.1637 Electron microscopy demonstrated the empty and abnormal-shape of LBs in the epidermis of WT/OVA, whereas ALCAM−/−/OVA LBs showed normal density and shape (Fig. 2C, insert C). In addition, ALCAM−/−/OVA showed higher LB count than WT/OVA (Fig. 2D). Quantitative real-time PCR analysis revealed that the levels of skin barrier genes including filaggrin, loricrin and involucrin were downregulated in WT/OVA, an effect that was mitigated by ALCAM-deficient mice (Fig. 2E). These findings suggest that ALCAM regulates the skin barrier function in AD.
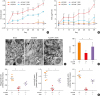 | Fig. 2ALCAM attenuates skin barrier disruption induced by epicutaneous OVA sensitization. (A) Time course of clinical score and (B) TEWL. (C) Electron micrographs of osmium tetroxide-postfixed skins show LBs in SG (×50,000, Scale bar = 0.5 μm; Insert C). (Left) Normal shaped LBs are present in sham mouse epidermis. (Middle) WT/OVA mouse epidermis shows severely abnormal LBs, (Right) Whereas LBs in ALCAM−/− mice display less abnormality (×120,000, Scale bar = 0.2 μm). (D) Bar graph represents the number of LBs in SG. (E) The mRNA expression of skin barrier genes (filaggrin, loricrin and involucrin).The data represent mean ± standard error of the mean.
ALCAM, activated leukocyte cell adhesion molecule; TEWL, transepidermal water loss; OVA, ovalbumin; PBS, phosphate-buffered saline; NS, not significant; WT, wild-type; LB, lamellar body; mRNA, messenger RNA; SG, stratum granulosum.
*P < 0.05 (WT/PBS vs. WT/OVA); †P < 0.05 (ALCAM−/−/PBS vs. ALCAM−/−/OVA); ‡P < 0.05 (WT/OVA vs. ALCAM−/−/OVA); §P < 0.05; ∥P < 0.01; ¶P < 0.001 (n = 5–7 mice/group).
|
ALCAM deficiency suppresses Th-2 inflammation
To investigate the contribution of ALCAM to Th2 inflammation, ALCAM−/− and WT mice were epicutaneously sensitized with OVA, and their inflammatory responses were compared. Infiltration of inflammatory cells and histological score were significantly increased in WT mice compared to those in ALCAM−/− mice following the EC OVA treatments (Fig. 3A). Moreover, WT/OVA mice manifest significantly increased total IgE accumulation (Fig. 3B) and heightened IL-4, IL-5, IL-13, interferon (IFN)-γ, IL-17A and IL-22 mRNA expression (Fig. 3C-H) compared with ALCAM−/−/OVA mice.
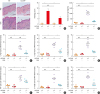 | Fig. 3ALCAM deficiency attenuates T helper-dominant skin inflammation induced by epicutaneous OVA sensitization. (A) Mouse skin biopsy specimen stained with hematoxylin and eosin (×200, Scale bar = 50 μm) and scored according to histological features. (B) Serum levels of total IgE measured by enzyme-linked immunosorbent assay. (C-H) Skin mRNA levels were assessed by quantitative real-time polymerase chain reaction; (C) IL-4, (D) IL-5, (E) IL-13, (F) IFN-γ, (G) IL-17A and (H) IL-22.The data represent mean ± standard error of the mean.
ALCAM, activated leukocyte cell adhesion molecule; NS, not significant; OVA, ovalbumin; PBS, phosphate-buffered saline; WT, wild-type; IgE, immunoglobulin E; IL, interleukin; IFN, interferon; mRNA, messenger RNA.
*P < 0.05; †P < 0.01; ‡P < 0.001 (n = 5–7 mice/group).
|
ALCAM deficiency suppresses CD4 T cell activation
Next, we investigated whether ALCAM deficiency affects Th2 cell activation in skin and skin-draining LNs by flow cytometry. After OVA sensitization, a lower accumulation of CD3+CD4+ T cells was detected in skin lesions (Fig. 4A) and skin draining LNs (Fig. 4C) of ALCAM-deficient mice than in those of WT mice. T cells were categorized as CD44loCD62Lhi (naïve), and CD44hiCD62Llo (effector memory).38 Interestingly, ALCAM-deficient mice had a reduced population of CD44hiCD62Llo cells in both skin and skin draining LNs relative to WT counterparts (Fig. 4A-D). These findings indicate that ALCAM mediates Th2-dominant inflammation via activation of CD4+ T cells in the skin and skin-draining LNs.
 | Fig. 4ALCAM deficiency suppresses CD4+ T cell activation. Cells from the (A and B) skin and (C and D) skin draining -LN(dLN)s were harvested and stained for CD3, CD4, CD44 and CD62L. Cells were gated on CD3+CD4+ cells and then on CD44 and CD62L. Naïve cells are identified as CD44loCD62Lhi and effector cells as CD4hiCD62Llo. Representative dot plots are presented. Graphs represent percentage of (B) CD3+CD4+CD44loCD62Lhi in skin and (D) CD3+CD4+CD4hiCD62Llo in dLNs.The data represent mean ± standard error of the mean.
ALCAM, activated leukocyte cell adhesion molecule; WT, wild-type; NS, not significant; OVA, ovalbumin; dLN, skin-draining lymph node.
*P < 0.05; †P < 0.01; ‡P < 0.001(n = 5–7 mice/group).
|
ALCAM deficiency suppresses skin inflammation and barrier disruption in the OXA- induced AD-like model
To further define the roles of ALCAM in allergic skin disease, we utilized AD-like mouse model that employed the OXA which could provoke an immune response. In these experiments, WT and ALCAM−/− mice were sensitized to and challenged with OXA, and the responses were evaluated. After OXA sensitization and challenge, WT mice manifest significantly increased clinical sore, TEWL and ear thickness compared to ALCAM−/− mice (Fig. 5A-C). Histological evaluations demonstrated enhanced epidermal thickness and epidermis inflammatory cell infiltration in comparisons of WT and ALCAM−/− mice (Fig. 5D and E). Next, we examined the total serum IgE level and tissue Th2 cytokines production. OXA-induced AD like WT mice showed increased total IgE accumulation (Fig. 5F) and heightened mRNA expression of IL-4 and IL-13 (Fig. 5G) compared to ALCAM−/− mice.
 | Fig. 5ALCAM deficiency attenuates murine AD-like skin lesion induced by repeated OXA application. (A-C) AD severity was assessed by (A) clinical score, (B) TEWL and (C) ear thickness. (D) Mouse ear skin biopsy specimen stained with hematoxylin and eosin (×100, bar = 200 μm) and (E) scored according to histological features. (F) Total IgE level in serum, as determined by enzyme-linked immunosorbent assay. The mRNA expression of (G) IL-4 and IL-13 in skin lesions.Data represent mean ± standard error of the mean.
ALCAM, activated leukocyte cell adhesion molecule; OXA, oxazolone; AD, atopic dermatitis; mRNA, messenger RNA; NS, not significant; WT, wild-type; TEWL, transepidermal water loss; IgE, immunoglobulin E; IL, interleukin, PBS, phosphate-buffered saline.
*P < 0.05 (WT/PBS vs. WT/OXA); †P < 0.05 (ALCAM−/−/PBS vs. ALCAM−/−/OXA); ‡P < 0.05 (WT/OXA vs. ALCAM−/−/OXA); §P < 0.05; ∥P < 0.01; ¶P < 0.001 (n = 4–6 mice/group).
|
The noted effects of ALCAM were not specific for allergens-induced inflammation because similar changes were noted in comparisons of skin tissues from ALCAM−/− and WT mice challenged with OXA.
DISCUSSION
In the present study, we addressed the involvement of ALCAM in AD pathogenesis using a murine model of OVA-induced AD. In our AD model, ALCAM levels were increased in serum relative to those in control mice. The serum ALCAM level was also more increased in AD patients that that in healthy controls; and was positively correlated with disease severity. Equivalent tendency of ALCAM alteration was described in our previous studies for allergic asthma and food allergy.2829 Although alterations in ALCAM level have been described in various human cancers,212439 alterations of ALCAM level in allergic disease has only recently been described, which is the first report providing evidence for the involvement of ALCAM in human AD.
Despite the repeated OVA exposure, ALCAM-deficient mice showed attenuated inflammatory responses and reduced production of Th2-type cytokines. We also observed a smaller population of CD3+CD4+ CD44hiCD62Llo T cells in the skin and skin-draining LNs of ALCAM-deficient AD mice. These results provide evidence that ALCAM is involved in AD pathogenesis.
A recent study showed that ALCAM is structurally similar to receptors for advanced glycation end products and can interact with S100B to activate nuclear factor κB signaling in delayed-type hypersensitivity diseases.4041 Although ALCAM may be associated with other inflammatory signaling pathways, our findings provide evidence that ALCAM–CD6 interactions contribute to both early and later stages of DC-induced T-cell activation and proliferation. It has also been reported that ALCAM–CD6 modulates the activation of the mitogen-activated protein kinase, extracellular signal-regulated kinase 1/2, p38 and c-Jun N-terminal kinase cascades,24 which further supports our conclusion that ALCAM contributes to the immune response.
As confirmed by our data, mouse models of AD induced by EC sensitization are characterized by epidermal thickening, CD4+ T cell infiltration, and up-regulation of the Th2 cytokines, IL-4, IL-5 and IL-13.34 Th2 cytokines regulate skin homeostasis via the signal transducer and activator of transcription-6.424344 Th2 inflammation also impairs the epidermal barrier, skin permeability, and cutaneous innate response.45 In AD mice, skin lesions showed reduced levels of skin barrier genes, filaggrin, loricrin, and involucrin and fewer lamellar bodies in the WT background than in the absence of ALCAM. Moreover, TEWL was less increased in ALCAM-deficient mice indicating the association of ALCAM with perturbation of the skin barrier. Hence, ALCAM may indirectly undermine the skin barrier function via modulation of the Th2 immune response.
In a psoriasis model, there is evidence that ALCAM affects T cell activation via Th17 cells.46 In the present study, ALCAM-deficient mice showed reduced expression of not only Th2 type cytokines but also IFN-γ, IL-17A and IL-22. As AD is a heterogeneous disease involving Th1, Th2 and Th17 cells, the role of ALCAM in the activation of various types of helper T cells and whether ALCAM expression levels differ in Th1, Th17 or Th2-dominated diseases need to be further elucidated.
In conclusion, the results presented here demonstrate that ALCAM can regulate Th2-type cytokine production and CD4+ T cell activation in AD-like skin lesions. In addition, our observations from clinical specimens suggest that ALCAM contributes to skin barrier dysfunction. These findings indicate that ALCAM mediates Th2-dominant immunological responses in AD pathogenesis and can be a potential target for disease treatment.
 XML Download
XML Download