

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Atopic dermatitis (AD) is a common chronic inflammatory skin disorder which is characterized by recurrent bacterial infections such as Staphylococcus aureus.12 This disease occurs mostly in infants and children with a high prevalence of approximately 20%, especially in developed countries.3 AD is a multifactorial complex disease that causes skin barrier dysfunction with different etiologies and prognoses.456 While some cases resolve spontaneously with time, others persist until adolescence and even develop into respiratory allergies such as asthma or allergic rhinitis.7 The severity of AD may be attributed to many environmental factors including exposure to inhalant and food allergens, air pollution, meteorological factors, and the microbiota of the skin.8 It is well known that S. aureus colonizes skin in 60%–100% of AD patients as compared to 5%–30% of healthy controls.910111213 Moreover, 10%–30% of S. aureus isolated from AD patients shows methicillin-resistant S. aureus (MRSA), although there are variations according to areas and the prevalence of MRSA infection is increasing.131415161718 In this review, we discuss S. aureus infection as an important determinant of AD severity19 and highlight its pathophysiologic roles, clinical implications and treatment in AD skin.
COLONIZATION OF AD SKIN BY S. AUREUS
AD patients colonized by S. aureus, especially MRSA, are related to previous hospitalization and use of topical calcineurin inhibitors in combination with topical steroids.14 It has been reported that early-life skin colonization of S. aureus may contribute to clinical AD onset during infancy.2021 S. aureus colonization could cause vicious cycles between S. aureus infection and AD exacerbation by inducing thymic stromal lymphopoietin (TSLP) and Th2/Th17-type inflammations.2223
Next generation sequencing techniques have advanced our understanding of changes in bacterial composition of the skin.2425 Commensal bacteria, such as Staphylococcus epidermidis and Staphylococcus hominis, modulate skin T-cell development, inhibit cutaneous inflammation and prevent skin infection by producing antimicrobial peptides (AMPs).122627 In contrast to the heterogenous communities of commensal bacteria in healthy skin, severe AD skin is dominantly colonized by S. aureus.123 In addition, S. aureus has greater predominance in patients with more severe AD. Nakatsuji et al.28 have reported that commensal bacteria, such as coagulase-negative Staphylococcus (CoNS), protects against S. aureus on normal healthy skin, but lack of commensal bacteria is associated with increased colonization of AD skin by S. aureus. Furthermore, they transplanted CoNS strains, such as S. hominis and S. epidermidis, on AD skin and found CoNS strains inhibited S. aureus. Recently, Shi et al.13 also studied staphylococcal colonization in a cohort of 339 AD patients and reported that microbial diversity was decreased on MRSA-colonized skin compared to methicillin-sensitive S. aureus (MSSA)-colonized skin. In addition, they reported that MRSA colonization of AD skin is associated with a more profound change in the composition of commensal bacteria than MSSA colonization.13 AD skin colonized by MRSA is associated with more severe skin inflammation compared to AD skin colonized with MSSA.26 Kennedy et al.29 found that bacterial community structures and diversity shifts over time and that infantile AD do not have noticeably dysbiotic communities before having AD. Of note, they also demonstrated that early colonization with commensal staphylococci at 2 months lowered the risk of developing AD at 1 year. However, further studies are required to confirm whether colonization by commensal bacteria could modify skin immunity and prevent development of AD.
WHY IS AD SKIN MORE SUSCEPTIBLE TO S. AUREUS COLONIZATION?
AD skin is characterized by chronic inflammation and recurrent bacterial infections by S. aureus.10 Multiple factors have been suggested as risk factors for S. aureus colonization of AD skin: the strength of S. aureus-corneocyte adhesion, deficiency of AMPs, decreased levels of filaggrin and filaggrin degradation products (FDPs), overexpressed Th2 cytokines, microbial dysbiosis, and altered lipid profiles (Fig. 1).4263031323334
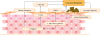 | Fig. 1
Staphylococcus aureus colonization on the skin of AD. Multiple factors, including skin barrier dysfunction, deficiency of AMPs, and decreased levels of filaggrin and FDPs, altered skin pH, overexpressed Th2/Th17 cytokines, microbial dysbiosis and altered lipid profiles, contribute to the colonization of S. aureus on AD skin.
AD, atopic dermatitis; FDP, filaggrin degradation product; AMP, antimicrobial peptide.
|
S. aureus initially attaches to the stratum corneum (SC), but the preferential adherence of S. aureus to AD skin has not been fully understood. Researchers have lately suggested that S. aureus isolated from AD skin binds with stronger affinity to deformed corneocytes, which are generated when FDP levels are reduced in SC.2535 Deformed corneocytes and the presence of fibronectin are likely to help S. aureus adhere to AD skin.25 Fibronectin-binding proteins and clumping factor B contribute to bacterial adherence to fibronectin, loricrin and cytokeratin 10.25
Cathelicidin (LL-37) and human beta-defensin (HBD)-3 are major AMPs that exhibit direct antimicrobial activities against S. aureus.2736 However, LL-37 and HBD-3 are down-regulated by interleukin (IL)-4 and IL-13, which are overexpressed on AD skin.3637 Decreased levels of these AMPs may be a risk factor for chronic colonization of S. aureus on AD skin.3839 Filaggrin and FDPs (natural moisturizing factors), such as urocanic acid and pyrrolidone carboxylic acid, play critical roles in maintaining skin pH and in inhibiting S. aureus growth on the skin.84041 It has also been reported that reduced levels of FDPs in SC are associated with more severe AD symptoms and induces strong adhesion of S. aureus to corneocytes of AD skin.35
Microbial dysbiosis induces pathologic bacterial infections such as S. aureus.2839 Skin commensal bacteria, such as S. epidermidis and S. hominis, are protective against S. aureus through production of AMPs.28 SC lipids, including free fatty acid, ceramides and sphingosine, play critical roles in the maintenance of skin barrier integrity and in preventing pathologic infections such as S. aureus. 43240 Berdyshev et al.30 recently found that lipid metabolisms and profiles for AD skin are altered by Th2 cytokines, which are highly expressed on AD skin.31 Therefore, aberrant lipid profiles could also exacerbate S. aureus colonization on AD skin.
IMPAIRMENT OF EPIDERMAL BARRIERS BY S. AUREUS INFECTION
S. aureus can cause both superficial and invasive infections of the skin, leading to bacteremia and sepsis.41 In addition, S. aureus colonizes the skin and produces virulence factors, including toxins, enzymes and other proteins, that contribute to inflammation and skin barrier dysfunction in AD.25 S. aureus on the epidermis can penetrate into the dermis depending on bacterial viability and protease activity in mice.25 Interestingly, this finding is correlated with increased expression of IL-4, IL-13, IL-17, IL-22 and TSLP, and with decreased expression of LL-37 as seen in AD.27 All S. aureus strains express superantigens such as staphylococcal enterotoxin (SE) A, SEB, SEC, SED and toxic shock syndrome toxin-1 (TSST-1) (Fig. 2). 4243 Highly abnormal and complex patterns of superantigens are found in more than 80% of S. aureus isolated from AD patients.19 Staphylococcal superantigens activate polyclonal T cells and subsequently cause T cell-mediated inflammation in AD lesions, by binding to the non-peptide groove of the major histocompatibility complex class II molecules on dendritic cells and T-cell receptors β chain on T cells without antigenic peptide presentation.25 In particular, SEB increases IL-31 expression and leads to the inhibition of keratinocyte differentiation and the suppression of filaggrin expression.4445 Superantigens also act as allergens and trigger immunoglobulin E (IgE) responses with histamine released from mast cells and basophils.25
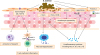 | Fig. 2The pathogenic effects of Staphylococcus aureus on atopic dermatitis. S. aureus expresses superantigens, such as SE and TSST-1, which activate basophils and cause T-cell-mediated inflammation. PSMα and PSMβ families stimulate the release of proinflammatory cytokines from keratinocytes. S. aureus α-toxin and δ-toxin induce keratinocyte cytotoxicity and mast cell degranulation, respectively. Protein A and lipoproteins of S. aureus lead to proinflammatory responses by activating TNFR-1 and TLR2 on keratinocytes, respectively.
SE, staphylococcal enterotoxin; TSST-1, toxic shock syndrome toxin-1, DC, dendritic cell; MHC II, major histocompatibility complex class II; TCR, T cell receptor; PSM, phenol-soluble modulins; TNFR-1, tumor necrosis factor receptor-1; TLR2, toll-like receptor 2; TSLP, thymic stromal lymphopoietin; IL, interleukin; KC, keratinocyte.
|
Short amphipathic peptides have been isolated by phenol extraction from S. aureus culture filtrates and are named phenol-soluble modulins (PSM) comprising the PSMα and PSMβ families along with δ-toxin. PSMs are secreted by the transporter Pmt (PSM transporter)46 and stimulates the release of proinflammatory cytokines such as IL-8 and IL-1β from keratinocytes.47 The δ-toxin is detected on the involved skin of AD patients and is known to cause mast cell degranulation.48
S. aureus α-toxin, also known as α-hemolysin, induces keratinocyte cytotoxicity in skin biopsy samples from AD patients.49 It has also been found that keratinocytes in AD is more susceptible to α-toxin due to reduced expression of filaggrin and sphingomyelinase by Th2 cytokines.49 Protein A and lipoproteins of S. aureus induce proinflammatory responses, such as TSLP and IL-8, by activating tumor necrosis factor receptor-1 and toll-like receptor 2 on keratinocytes, respectively.222550 Taken together, S. aureus exacerbates inflammation on AD skin through release of superantigens, PSMs, toxins and lipoproteins that affect keratinocytes and immune cells.
WHY IS IN AD SKIN INFECTION WITH S. AUREUS IMPORTANT IN CLINICAL PRACTICE?
There has been increasing evidence that S. aureus plays pivotal roles in the exacerbation of AD as well as infectious complications such as impetigo, cellulitis, abscesses and invasive infections.51 Colonization of S. aureus leads to more severe disease as well as exacerbation of AD.92021 The density of S. aureus on both affected and unaffected skin is associated with increased severity of the Scoring Atopic Dermatitis index.52 A decreased diversity of the skin microbiota and increased abundance of S. aureus were observed during flares of AD.53 Additionally, MRSA colonization was significantly associated with the misuse of antibiotics and previous hospitalization.11418 The reason for this relationship is believed to be that S. aureus produces toxins, proinflammatory cytokines and proteases that cause dysregulation of skin homeostasis by affecting keratinocytes and various immune cells in AD skin.5455
S. aureus colonization is strongly linked to more severe barrier dysfunction in terms of the integrity of the SC, permeability and skin hydration even on non-lesional AD skin.56 For example, AD patients colonized with S. aureus show higher transepidermal water loss compared to noncolonized controls or those patients who are not colonized.56 Interestingly, different clonal complex (CC) types are found between S. aureus strains isolated from patients with AD and those from unaffected carriers. 5758 Clausen et al.59 reported that temporal changes in these CC types are associated with the clinical course of AD. Changes in CC type during follow-up were related to increased severity of AD, while patients carrying the same CC type showed a reduction in severity scores.59 They also demonstrated that skin pH was higher in patients colonized with S. aureus than in those not, indicating a relationship between skin pH and imbalance of the skin micriobiome.59
S. aureus colonization is associated with greater Th2 polarization, allergen sensitization and tissue damage compared to non-colonized subjects.56 Therefore, it is postulated that cutaneous infection with S. aureus contributes to the development of atopic march by enhancing chronic skin inflammation and allergen sensitization.7 Indeed, a recent study showed that enterotoxin-producing S. aureus not only resulted in systemic Th2 responses, but also exaggerated allergen-induced lung inflammation and airway hyperresponsiveness through an IL-17A dependent mechanism in a mouse model.60 It has also been suggested that S. aureus colonization in AD patients is associated with food allergy.6162 Jones et al.61 recently reported that there was an association between S. aureus colonization and food allergy including peanut, egg white and cow's milk in children with AD. Moreover, they found peanut specific IgE levels were significantly higher in patients with MRSA colonization compared to those with MSSA, suggesting a stronger association between MRSA colonization and peanut allergy. However, further studies are necessary to elucidate how S. aureus colonization affects the atopic march and food allergy in children with AD.
HOW TO MANAGE S. AUREUS SKIN INFECTION IN AD PATIENTS
Skin colonization by S. aureus, particularly MRSA, is one of the major challenges commonly encountered in the management of AD.1 Impetigo can be treated with either topical mupirocin or topical retapamulin for 5 days.63 Oral antibiotics, such as antistaphylococcal penicillins or first-generation cephalosporins, are used for 7 days when MSSA infection is of concern.6364 For the treatment of MRSA skin and soft tissue infection, the Infectious Diseases Society of America recommends doxycycline, clindamycin or sulfamethoxazole-trimethoprim, while other guidelines suggest vancomycin, teicoplanin, glycopeptides, daptomycin, telavancin, linezolid or tigecycline as well.6365 However, antibiotics may be less effective due to the development of antibiotic-resistant strains, recolonization and their negative impact on the beneficial commensal microbes.2
Recent studies have reported that patients with AD can obtain benefit from the use of bleach (sodium hypochlorite) bath.66 Bleach bath seems to improve AD through complex antimicrobial, anti-inflammatory, and antipruritic effects on the skin.67 However, it is possible that the nonselective antimicrobial effect of bleach could affect both pathogenic and commensal microbes.67 Moreover, a recent meta-analysis failed to show additional benefits of bleach bath compared to water bath alone, suggesting the need for further research in order to establish the effects and long-term safety of this therapeutic strategy.68
Interestingly, microbiome manipulation with topically applied commensals has been introduced to eradicate S. aureus from AD skin.28 The application of CoNS strains with antimicrobial activity to human subjects with AD reduced colonization by S. aureus, indicating the important role of skin dysbiosis in the pathogenesis of AD.28 Additionally, several studies, including the use of monoclonal antibodies against virulent toxins, vaccines and recombinant phage endolysin, have been conducted in various stages.6970
CONCLUSION
S. aureus commonly colonizes the skin of AD patients and contribute to the development and exacerbation of AD. AD skin is susceptible to S. aureus colonization due to deficiency of AMPs, decreased levels of filaggrin and FDPs, and microbial dysbiosis. S. aureus not only impairs skin barrier function directly, but also up-regulates proinflammatory cytokines leading to Th2/Th17 polarizations and skin inflammation. Bleach bath and antibiotics have been used to reduce the burden of S. aureus. Moreover, efforts to restore skin microbial dysbiosis have been tried. Further advances in our understanding of the role of S. aureus in the pathophysiology of AD will allow us to manage skin symptoms more effectively in AD patients .
 XML Download
XML Download