

 PDF
PDF Citation
Citation Print
Print



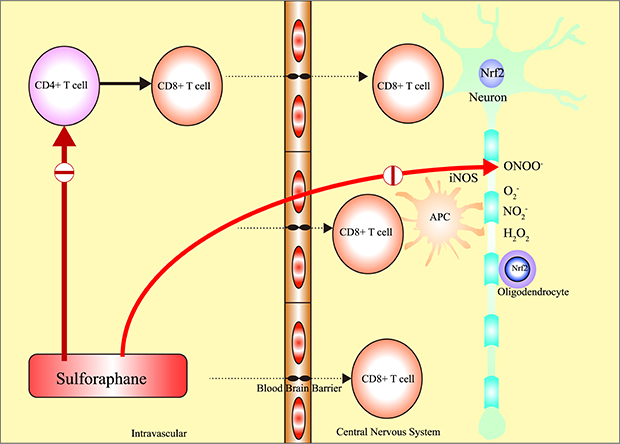
INTRODUCTION
Multiple sclerosis (MS) is an immune-associated inflammatory disorder of the central nervous system (CNS) and effects more than 2 million patients worldwide.1234 The etiology of MS has been characterized by crossing of the blood–brain barrier (BBB) by T-lymphocytes and demyelination, reactive gliosis, and axonal degeneration by activated intravascular autoimmune T-cells.134 In addition, previous studies have shown that MS is related with environmental influences, altered genetic factors, vitamin D deficiency, and relevance to other autoimmune disorders.45678
Accordingly, most disease-modifying therapies (DMTs) for the treatment of MS have been adapted to the autoimmune related mechanism. However, DMT drugs have limitations, such as expensive cost, several fatal side effects such as progressive multifocal leukoencephalopathy (PML), and unsatisfactory treatment response.91011 Therefore, further development of new therapeutics has been greatly required for MS.
Recently, some documents have reported that oxidative stress is associated with the onset of MS.12131415 The overproduction of reactive oxygen species (ROS) and impairment of the anti-oxidative system result in increased oxidative stress, which activates pro-inflammatory cytokines, which contribute to the development and progression underlying the pathogenesis of MS.16
Sulforaphane (SFN; 1-isothiocyanate-4-methyl sulfonyl butane) is an anti-oxidant phytochemical derived from cruciferous vegetables, such as broccoli. Through its anti-oxidative effect, it exerts neuroprotective effects via nuclear factor erythroid 2-related factor 2–Kelch-like-ECH-associated protein 1–antioxidant response element signaling pathway in other diseases, such as Alzheimer's disease, Parkinson's disease, tumor and heart problems.171819202122232425
Herein, we studied the neuroprotective effects of SFN using a mice model of experimental autoimmune encephalomyelitis (EAE) which is usually used in studies of MS because EAE mice model is characterized by perivascular infiltration of inflammatory cells and followed by demyelination in the brain and spinal cord via CD4+ T-cell–mediated autoimmune cascade.262728293031
The purpose of this study was to reveal whether SFN has protective effect against autoimmune-associated demyelination through its anti-inflammatory and anti-oxidative effects in EAE mice.
METHODS
Induction of EAE mice and analysis
We used a C57BL/6 mice model that had properties of auto-immunization against the myelin oligodendrocyte glycoprotein (MOG35–55) peptide that is widely applied to induce EAE in animals.3032 All procedures were conducted by following the Institutional Animal Care and Use Committee (IACUC) guidelines; all operations were carried out under sodium pentobarbital anesthesia, and all procedures were done to minimize agony. During the week before and during the experimental processing, the mice were cared for in cages with room ambient temperature of 23°C (± 3°C), durable indoor humidity, and the alternation of day and night.
Twenty C57BL/6 female mice of 9-week-old age (Central Lab. Animal Inc., Seoul, Korea) were used in this study. To experiment the effects of SFN, mice were randomly assigned to two groups:1) C57BL/6 mice treated with 50 mg/kg/day SFN (n = 10) and 2) C57BL/6 mice treated with a vehicle (phosphate-buffered saline [PBS], n = 10). Before EAE induction, SFN or PBS was administrated daily for 14 days in each group. All C57BL/6 mice were auto-immunized against the MOG35-55 (MEVGWYRSPFSRVVHLYRNGK) peptide in incomplete Freund's adjuvant supplemented with Mycobacterium tuberculosis (EAE induction Hooke kits™, catalog No. EK-2110; Hooke Laboratories, Inc., Lawrence, MA, USA) following the manufacturer's instructions. Briefly, MOG (200 µg) was subcutaneously injected and pertussis toxin (200 ng) was intraperitoneally injected once in each mouse as previously described.3334 After the autoimmunization, further doses of SFN (50 mg/kg) or PBS were administered orally once per day. Subsequently, SFN or PBS was administrated daily for 28 days in each group, then spinal cords were removed for biochemical or immunohistochemical analyses.
After EAE induction, all mice were checked daily, and their behavioral states were documented. EAE mice showed decreased muscle tone, which is characterized by progression from the tail upward through the body.353637 A six-point scale (0–5) was used to rate and monitor the degree of the symptoms, with a score of 1 which means loss of tail tonicity, 4 indicating quadriplegia, and 5 signifying death, as previously described.383940 An investigator blinded to the groupings used the following grading scores: 0, no clinical symptom; 0.5, piloerection; 1, mild loss of tail tone; 1.5, complete loss of tail tone; 2, mild hind-limb paresis; 3, complete bilateral hind-limb paralysis; 3.5, complete bilateral hind limb paralysis and unilateral fore-limb paresis; 4, total paralysis of fore and hind limbs; 5, death. The symptom scores are expressed in mean ± standard error (SE).
Tissue extraction
All animals were euthanized on the 14th day after EAE induction by intra-cardiac perfusion using 4% paraformaldehyde. First, mice were given anesthetics by intraperitoneal injection, and thoraco-abdominal incision was performed to disclose the heart. Then, a 24-gauge needle was inserted into the left ventricle, and 25–50 mL of cold saline solution mixed with 1 U/mL heparin sodium (Green Cross Co., Fukuoka, Japan) was injected. The blood was excreted freely through an incision in the right atrium. Thereafter, a similar amount of 4% paraformaldehyde (pH 7.4) was injected with the same rate of perfusion. After perfusion, brain and spinal cord specimens were fastened with 4% (w/v) paraformaldehyde in PBS buffer for a night at 4°C. The temperature was set to 4°C and high sucrose concentration (30%) was used to dehydrate and sink the samples. To make frozen blocks, the specimens were placed in O.C.T. compound (Sakura Finetek USA, Inc., Torrance, CA, USA) and frozen with dry ice.
Immunohistochemistry
The cryostat was used to cut the histological slides into 10-µm thick sections and slides were collected from the cervical to the thoracic spine and stained with hematoxylin and eosin (H&E) and luxol fast blue (LFB) to analyze inflammation and demyelination. The sections were washed with distilled water (DW) for 5 minutes and stained with hematoxylin solution (Mayer's Hematoxylin solution; Sigma-Aldrich, St. Louis, MO, USA) for 10 minutes. After being washed with DW, the slides were stained with Eosin Y solution (Sigma-Aldrich) for 3 minutes. Also, for LFB staining, the sections were washed with DW and then incubated for a night at 60°C in LFB (concentration 0.1%) (Solvent Blue 38; Sigma-Aldrich). In all sections, light microscopic analysis was used to evaluate the slides.
Immunoblotting assay
Fourteen days after immunization, the mice were perfused with PBS while under deep anesthesia, and tissues of brain and spinal cord were collected. Spinal cord tissues of each mouse were made homogeneous in ice-cold radio immunoprecipitation assay buffer (catalog No. R2002; Biosesang, Seongnam, Korea) with protease inhibitor and phosphatase inhibitor cocktails (catalog No. 01906845001 and 11697498001, Roche Diagnostics Corporation, Indianapolis, IN, USA). Centrifugation was used to collect the supernatants at 14,000 rpm and 4°C for 25 minutes. The samples were denatured in sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) loading buffer (Biosesang, cat# S2002) for 5 minutes at 100°C, put into 15% gel for separation by SDS-PAGE, and then absorbed into nitrocellulose membranes with a Trans-Blot Cell system (Bio-Rad Laboratories, Inc., Hercules, CA, USA). And, the membranes were sealed off for one hour at room ambient temperature with 5% skim milk (Duchefa Biochemie B.V., Haarlem, Netherlands) in Tris-buffered saline (20 mM Tris, 150 mM NaCl [pH, 7.5]) with 0.1% Tween 20. The membranes were incubated overnight at 4°C with the hereinafter antibodies: anti-MOG antibody (1:1,000; catalog No. ab109746, Abcam plc, Cambridge, UK), anti-myelin basic protein (MBP) antibody (1:2,000, catalog No. ab40390, Abcam plc), anti-glial fibrillary acidic protein (GFAP) antibody (1:10,000, catalog No. ab7260, Abcam plc), anti-iNOS antibody (1:500, sc-7271; Santa Cruz Biotechnology, Inc., Dallas, TX, USA), anti-CD68 antibody (1:200, sc-20060, Santa Cruz Biotechnology, Inc.), and anti-CD4 antibody (1:200, sc-1140, Santa Cruz Biotechnology, Inc.); anti-glyceraldehyde 3-phosphate dehydrogenase (1:3,000, catalog No. 2118, Cell Signaling Technology, Inc., Denver, MA, USA) was used as a loading control.
Statistical analysis
The symptoms score was expressed as mean ± SE. SPSS 21.0 (IBM Corp., Armonk, NY, USA) was used for statistical analysis. Statistical analyses were conducted using Wilk-Shapiro test for normality and Mann-Whitney U test. P values less than 0.05 were considered statistically significant.
Ethics statement
All of the procedures were performed in accordance with the IACUC guidelines and were approved by IACUC (permit No. 2015-00026) of the Chung-Ang University for the care and use. Written informed consent was not used because of the nature of the study design, which utilized animal model.
RESULTS
Clinical score
In this experimental study, we analyzed the clinical symptom scales in individual EAE groups daily to examine the functional loss and alterations. After EAE mice induction by auto-immunization against MOG35–55 peptides, the EAE symptom scales increased steadily in each group, and statistical differences were significantly identified between the groups at the 13th and 14th day after immunization. The SFN-administered mice had lower symptom scores than the untreated mice, which were statistically significant at the 13th day (1.30 ± 0.15 vs. 1.90 ± 0.18; P = 0.043) and 14th day (1.80 ± 0.13 vs. 2.75 ± 0.17; P = 0.003) (Fig. 1). These results demonstrated that the SFN administration alleviated the clinical symptoms and suppressed disease progression in the EAE mouse model.
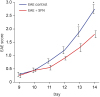 | Fig. 1Results of behavioral study using clincal symptom scores. After autoimmunization with MOG35–55, the EAE symptom scores gradually increased in each group, and significant differences were identified among the groups at the 13th and 14th day after immunization. The SFN-administered mice had lower symptom scores than the untreated mice, statistically significant at the 13th day and 14th day. The results are expressed as mean ± standard error.
MOG = myelin oligodendrocyte glycoprotein, EAE = experimental autoimmune encephalomyelitis, SFN = sulforaphane.
*P < 0.05.
|
Biochemical results
Using immunohistochemistry, we examined the spinal cord specimens for inflammatory cell infiltration and demyelination to identify whether the pathological and functional alterations of the EAE mice were influenced by the oral administration of SFN. To identify the effectiveness of SFN on inflammatory process and demyelinating changes in the white matter, we had stained the different levels of spinal cord sections. After identifying specimens, those from cervical spinal cords were not adequate for histologic analysis. Therefore, we analyzed only specimens from thoracic spinal cords with H&E (Fig. 2) and LFB staining (Fig. 3).
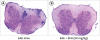 | Fig. 2H&E stained spinal cords in the EAE mice. (A) The untreated EAE mice revealed increased intensity of H&E staining in white matter of spinal cord compared with (B) SFN treated EAE mice.
H&E = hematoxylin and eosin, EAE = experimental autoimmune encephalomyelitis, SFN = sulforaphane.
|
 | Fig. 3Luxol fast blue stained sections of spinal cords in the EAE mice. (A) The untreated EAE mice revealed prominent demyelination in white matter of spinal cord compared with (B) SFN treated EAE mice.
EAE = experimental autoimmune encephalomyelitis, SFN = sulforaphane.
|
SFN administration remarkably influenced the inflammatory activity and myelination in the spinal cords of the EAE mice. The SFN-treated EAE mice revealed decreased inflammatory infiltration in the H&E staining (Fig. 2) and relatively preserved myelination in the LFB staining (Fig. 3) of the spinal cord compared with control EAE mice.
On the 4′,6-diamidino-2-phenylindole (DAPI)-MBP double staining, the untreated-EAE mice showed more prominent DAPI-positive and MBP-negative findings of spinal cords compared with the SFN-treated EAE mice (Fig. 4). These results demonstrated that SFN treatment inhibited spinal cord demyelination following the infiltration of inflammatory cells in the EAE mice.
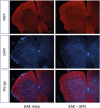 | Fig. 4The DAPI-MBP double staining. The untreated-EAE mice showed more prominent DAPI-positive and MBP-negative findings of spinal cords compared with the SFN-treated EAE mice. These results demonstrated that SFN treatment inhibited spinal cord demyelination following the infiltration of inflammatory cells in the EAE mice.
DAPI = 4′,6-diamidino-2-phenylindole, MBP = myelin basic protein, SFN = sulforaphane, EAE = experimental autoimmune encephalomyelitis.
|
Furthermore, we evaluated CNS inflammation by identifying the presence of activated microglial cells (CD68), GFAP and CD4 T-cell; in addition, we assessed iNOS expressions in the EAE mice to identify the changes related to antioxidant processes. Collectively, the results of our study confirmed the anti-inflammatory and antioxidant effects of SFN.
Compared with the untreated EAE mice, the SFN-treated EAE mice showed decreased immunoreactivity to CD4 T-cell (Fig. 5), GFAP and CD68 (Fig. 6). These results suggest that SFN suppressed the autoimmune cascade reactions in the EAE mice by the inhibition of T-cell activation.
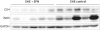 | Fig. 5Compared with the untreated EAE mice, the SFN-treated EAE mice showed decreased immunoreactivity to CD4 and iNOS.
EAE = experimental autoimmune encephalomyelitis, iNOS = inducible nitric oxide synthase, GAPDH = glyceraldehyde 3-phosphate dehydrogenase, SFN = sulforaphane.
|
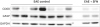 | Fig. 6Compared with the untreated experimental EAE mice, the SFN-treated EAE mice showed decreased immunoreactivity to GFAP and CD68.
EAE = experimental autoimmune encephalomyelitis, GFAP = glial fibrillary acidic protein, GAPDH = glyceraldehyde 3-phosphate dehydrogenase, SFN = sulforaphane.
|
To evaluate the anti-oxidative effects of SFN on ROS activity in mice with EAE, we measured iNOS expression using WB. SFN effectively inhibited the iNOS up-regulation induced by oxidative stress in the EAE mice and this result revealed the inhibition of stimulated nitric oxide (NO) production followed by the anti-oxidative effect of SFN in the mice with EAE (Fig. 5).
DISCUSSION
MS is an immune-associated demyelinating disease involving brain, spinal cord and optic nerve that results in serious disability. Recently, significant progresses have been reported toward revealing the immune-related pathogenesis of MS and the effectiveness of DMTs in preventing relapse.414243 Although many DMT drugs are reported to be effective in patients with MS, current DMT drugs have many limitations, such as highly expensive cost and serious adverse effects, such as cardiac problems, PML, and teratogenic effect.9101115444546474849 Therefore, there has been reasonable need for safe, effective, and less expensive therapeutic options.
Recent experimental studies have implicated ROS focusing on superoxide, NO, and their intermediates, as contributors to several mechanisms underlying the pathogenesis of MS and EAE.1213141550 ROS has been reported to promote leukocyte traffic across the BBB. Furthermore, infiltrated leukocytes produce ROS that induce myelin breakdown by macrophages, oligodendrocyte damage, and axonal and neuronal injury. In addition, reactive microglia generate peroxynitrite, a principal mediator of the oxidative stress and excitotoxicity that drive neurodegenerative processes in MS.
SFN is derived from cruciferous vegetables, such as broccoli and cabbage. In patients with several diseases and animal models of Alzheimer's disease and Parkinson's disease, SFN showed anti-inflammatory and anti-oxidative effects.182122232425
In this experimental study, we revealed anti-inflammatory effects of SFN in EAE mice and found that it reduced the aggravation of the clinical symptoms. Also, SFN showed anti-oxidative effects evidenced by less NO production and inhibited the infiltration of abnormal inflammatory cells, and demyelination in the spinal cord of EAE mice. EAE model is considered similar with MS in terms of the abnormal activation of T-cells, and followed by inflammatory cells into the CNS, which results in the autoimmune-associated neurological disease. Thus, the results of this study suggest that SFN would be valuable in the treatment or prevention of MS.
Therefore, considering the recent advantages of botanical drugs, if SFN is developed as a new drug, there will be some advantages.5152 Actually, SFN is relatively inexpensive because SFN is abundant in easily available vegetables, such as broccoli, and it can be expected to have much fewer side effects than DMTs, making it the safer option. Furthermore, the combination of SFN and other DMTs may also have additive effects for the patients with MS.53
Our study identified the therapeutic efficacy of SFN administration in EAE mice, thus SFN may be highly feasible as a strategic therapeutic for patients with MS, and this experimental study can be the potential proposal to plan the clinical research on treating MS using SFN.
Of course, there were some limitations in this study. The sample size of EAE mice was not enough to show the expected therapeutic effect between-group difference. And the grouping of EAE mice was not systematized to identify the treatment period and therapeutic dosage of SFN. Additionally, further biochemical analysis using more specific cytokines and metabolites is necessary.
The therapeutic effects of SFN in EAE mice have not been reported to our knowledge. In our study, the SFN treatment showed anti-inflammatory and anti-oxidative effects in EAE mice, therefore the SFN could be a new therapeutic or nutritional supplement for patients with MS.
 XML Download
XML Download