

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Dear Editor:
Acne is inflammatory disorder of the pilosebaceous unit. Its mechanism is complex, occurring as a result of hyperkeratinization of the pilosebaceous ducts, colonization of Propionibacterium acnes, and perifollicular inflammation with an imbalance of sebum production1. Insulin-like growth factor (IGF)-1 and peroxisome proliferator-activated receptor (PPAR) regulate the growth and differentiation of sebocytes and sebum secretion. Hyperglycemic diets induce IGF-1 production and may aggravate acne2. Eupatilin [2-(3,4-dimethoxyphenyl)-5,7-dihydroxy-6-methoxychromen-4-one] is the main flavonoid of the Artemisia species. Eupatilin exerts various effects including anti-apoptotic, cytoprotective, antioxidant, and anti-inflammatory nature on many different cell lines3. Eupatilin demonstrates antioxidant activity by suppressing reactive oxygen species and anti-inflammatory activity by inhibiting 5-lipoxygenase4. In addition, eupatilin downregulates the production of inflammatory cytokines such as tumor necrosis factor (TNF)-α, interleukin (IL)-4, IL-6, and IL-18 through nuclear factor kappa B (NF-κB) in RBL-2H3 cells345. Eupatilin has been used in an attempt to address inflammatory diseases, but its therapeutic effect on acne and related mechanisms remain unclear. In this study, we aimed to investigate the therapeutic effect of eupatilin on IGF-1-induced inflammation and lipogenesis using human sebocytes.
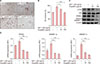
To examine the effects of eupatilin in vitro, we used human SZ95 sebocytes (Supplementary Material). The effects of eupatilin on the proliferation of cultured SZ95 sebocytes were determined with various doses of eupatilin. In MTT assay, the proliferation of SZ95 sebocytes was sequentially decreased according to the concentration (10, 20, 50, or 100 µM of eupatilin) (Supplementary Material). Thus, we determined experimental concentration as 10- and 100-µM of eupatilin. Next, we stained an intracellular lipid droplet formation using Oil Red O to investigate the effects of eupatilin on the lipid synthesis of sebocytes. When SZ95 sebocytes were treated with eupatilin, lipid accumulation in the cytoplasm was significantly reduced (Fig. 1A, B). To elucidate how eupatilin suppressed IGF-1-induced lipogenesis of SZ95 sebocytes, the effects of eupatilin treatment on the IGF-1-induced expression levels of phosphorylated Akt and lipogenesis-related transcription factors (PPARγ and mature sterol regulatory element-binding protein [SREBP]-1) were measured. In comparison with no treatment, 100 µg/ml of eupatilin significantly reduced the protein levels of phosphorylated Akt, PPARγ, and mature SREBP-1 of the sebocytes, which were increased by IGF-1 pretreatment (Fig. 1C). Likewise, eupatilin also significantly downregulated the mRNA expression level of PPARγ, SREBP-1a, and SREBP-1c of the sebocytes (Fig. 1D). These results suggest that eupatilin has an inhibitory effect on IGF-I-induced lipogenesis of sebocytes through suppression of the phosphorylation of Akt, PPARγ, and mature SREBP-1.
To further investigate the anti-inflammatory effects of eupatilin on sebocytes, we analyzed the IGF-1-induced proinflammatory cytokines of sebocytes. In the immunofluorescence study, treatment with eupatilin inhibited the translocation of NF-κB p65 induced by 50 ng/ml of IGF-1 (Fig. 2A). This eupatilin treatment also significantly downregulated the mRNA expression levels of (pro)inflammatory cytokines such as TNF-α, IL-6, and IL-8 (Fig. 2B). In this study, we first showed that the proper concentrations of eupatilin (10 µg/ml and 100 µg/ml) had potent anti-IGF-I effects on SZ95 sebocytes, as follows: 1) eupatilin suppressed the lipogenesis of sebocytes by interfering with the expression of phosphorylated Akt, PPARγ, and SREBP-1 induced by IGF-1; and 2) eupatilin reduced the inflammatory response of sebocytes by inhibiting NF-κB activation, with IGF-1 able to induce the inflammatory cytokine expression of sebocytes.
SREBP-1 is a major transcription factor that regulates cholesterol/fatty acid metabolism6. Smith et al.7 reported that IGF-1 induced SREBP-1. This activation occurred via a phosphoinositide 3-kinase (PI3K)/Akt pathway. They also showed that IGF-I transmits its lipogenic signal in sebocytes via an Akt pathway. Meanwhile, PPARγ is a potential modulator of lipid production in human sebocytes8. PPARs are regulators of lipogenesis and differentiation of keratinocytes as well as sebocytes and have three isoforms (i.e., α, δ, and γ). Notably, PPARγ is important for sebaceous gland development and function9. Interestingly, in the present study, the Oil Red O stain showed that eupatilin suppressed lipid synthesis by sebocytes. Although eupatilin from Artemisia plants has been reported as a selective PPARα agonist10, in the present study, the proper concentrations of eupatilin (10 µg/ml and 100 µg/ml) inhibited the IGF-1-induced SREBP-1 and PPARγ expression of sebocytes. In addition, eupatilin inhibited the phosphorylation of Akt. This inhibitory effect of eupatilin on IGF-1-induced sebocytes was confirmed by the reduced mRNA expression level of PPARγ, SREBP-1a, and SREPB-1c. Moreover, we found that eupatiln suppressed the mRNA expression of TNF-α, IL-6, and IL-8 of sebocytes at the transcription level. Likewise, eupatilin also inhibited IGF-1-induced nuclear translocation of NF-κB p65 of sebocytes. This study demonstrate that eupatilin strongly downregulated IGF-I-induced inflammation and lipogenesis of SZ95 sebocytes. Therefore, our results suggest that eupatilin could be a promising candidate for the management of acne.

 XML Download
XML Download