

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
In recent years, the use of medicinal plants has received much attention [1]. In this regard, genistein as an angiogenesis inhibitor and a phytoestrogen found in soy and some plant species has many beneficial properties for health [2]. Approximately, 99% of genistein is found in soybeans combined with glucose molecules [3]. Some therapeutic effects of genistein have been reported on the inhibition of some diseases. It seems that, due to isoflavone administration, the blood glucose level is reduced in diabetic rats [45]. Moreover, genistein has antiinflammatory and antioxidant activity [6]. Nicotine is an alkaloid formed by pyridine and pyrrolidine loops found in the tobacco plant [7]. Oxidative stress and an increase in lipid oxidase production occur following nicotine injection, which may lead to irreversible damage of the cellular membrane. The increased level of reactive oxygen species (ROS) causes oxidative stress and induces DNA breakage [8]. Smoking of tobacco, particularly cigarettes, has a negative effect on the pancreases. Nicotine can induce oxidative stress in the animal pancreas and this was along with inflammation and improved interleukin 6 secretion in the pancreas [9]. Expression of inducible nitrite oxide synthase occurs following the advance of pancreatic cancer as well as in inflamed tissues [10]. Genistein is active in many biochemical pathways and inhibits intracellular enzymes [11]. Increased ROS causes DNA failure and impairs biologic membranes by inducing oxidative stress [12]. According to genistein effects and since no article has yet been reported on the effect of genistein in contradiction of nicotine injuries, the present study was designed to evaluate the effect of genistein on disorders and damage induced by nicotine in animal samples.
Materials and Methods
Animals
In this experimental study, 48 BALB/c male mice (25–30 g) were purchased from Pasteur Institute. The animals were housed in standard cages and control conditions at 23±2℃ and exposed to a 12-hour light/dark cycle, in animal care center of Kermanshah Medical Sciences University. All investigations conformed to the ethical and humane principles of research and were approved by the Ethics Committee of Kermanshah University of Medical Sciences (No. 1395.600) [8].
Protocol and treatments
The animals were randomly separated into six groups (n=8). Group 1 (normal control or saline group) received 0.9% normal saline. Group 2 (nicotine control group) was induced by nicotine (2.5 mg/kg). Groups 3 and 4 (genistein groups) were given 25 and 50 mg/kg genistein, respectively. Mice in groups 5 and 6 received genistein (25 and 50 mg/kg)+nicotine. Nicotine through inter peritoneal injection administrated for 4 weeks every day at 10 a.m. in order to induce pancreas damage. Mice with genistein were administrated as follows: On days 1–30, genistein once daily, intraperitoneally injecting. Mice with nicotine+genistein were administrated as follows: on days 1–30, genistein once daily+nicotine. The same volume of saline was administered to all groups [813].
Dissection, pancreas weight and collection of blood serum
Animals were anesthetized with chloroform and dissected, followed by taking blood samples from their right ventricle by a cardiac puncture. The samples were incubated for 15 minutes at 37℃ to clot. Then, the clotted blood was centrifuged for 15 minutes at 3,000 rpm to acquire the serum. In order to measure the weight of the pancreas, pancreas extraction and weighed as follows. The thoracic cavity was opened. Skin of the mouse between the two hind legs was cut and the xiphoid was removed. The stomach was located on the left side of the mouse. It was gently (so as to avoid tearing) separated the pancreas from the stomach and duodenum by using two forceps. When detaching the pancreas from the stomach and intestines, it is very important that the forceps are used gently to guide the pancreas tissue away from the organs and to not crush or tear the pancreas with the forceps. The pancreas was continued to separate from the small intestine jejunum and ileum sections, and lastly from the caecum of the large intestine. At the caecum, the forceps were repositioned and continued separation of the pancreas along the remaining colon towards the rectum. Locate the pancreas and attached spleen. The pancreas slide towards the right side of the mouse. The remaining connections between the pancreas and thoracic cavity was separated with the forceps to fully detach the pancreas and adjoined spleen. The pancreas was removed and spread it out for examination. The spleen was attached to the pancreas for identification purposes. Then, the nearby fat, intrapancreatic bile duct and capsule were removed and then weighed on a microbalance sensitive up to 0.001 mg (Precisa, Zurich, Switzerland) and average weights were calculated and recorded (Fig. 1) [14].
Nitrite oxide assay
Nitrite oxide measurement was made using the Griess assay. After de-freezing the serum samples, supernatant (500 µl) was deproteinized, 200 µl supernatant was taken, and then 200 µl vanadium chloride, 60 µl N-(1-naphthyl) ethylenediamine dihydrochloride, and 60 µl sulfonamide solutions were added. Increasing concentrations of sodium nitrite (5, 10, 25, 50, 75, and 100 µM) were used for the standard curve samples and was measured using enzyme-linked immunosorbent assay (ELISA) reader (Hyperion, Miami, FL, USA) at a wavelength of 450 nm according to manufacturer's protocol [12].
Histological examinations
The pancreas was fixed in 10% formalin and then cut transversely into three parts. The central part was immersed in 70% alcohol, dehydrated in ascending concentrations of ethanol, cleared in xylene, and then embedded in soft paraffin and sectioned (5-µm-thick) using a microtome (EC350-2, Leica, Nussloch, Germany). Using the hematoxylin and eosin staining, the morphometric assays were tested under an Olympus BX-51T-32E01 microscope (Tokyo, Japan) linked to a DP12 camera and Olysia Biosoftware (Olympus) [14].
Morphometric study
To measure the number and thickness of pancreatic islets, 10 scratches were stained from each section. Afterward, five fields in the 100× magnification of microscope were randomly selected. Followed by calculating the number of islets, their mean was attained. To quantify the mean diameter of the islets, five of them were selected from each lam. Then, the largest and smallest thickness of each islet was determined in micrometer and by the following method, the mean diameter of each islet was acquired.
, where MD, mean diameter; S, small diameter of the islet; and L, large diameter of islet [15].

Biochemical assay
Insulin was measured using the ELISA method by Monobind kit (Sigma, St. Louis, MO, USA) according to manufacturer's instructions. Plates were read by ELISA reader at 450 nm [16].
Ferric reducing ability of plasma method
Ferric reducing ability of plasma (FRAP) method was used to measure the total antioxidant capacity of the serum. The plasma capability to restore ferric ions was measured in this method. This process required a great quantity of FeIII. When the compound of FeIII-TPTZ was reverted to FeII, a blue color formed and absorption occurred at the maximum wavelength of 600 nm. The FRAP substance was consisted of 30 ml of acetate buffer (Sigma) and 1.5 ml ferric chloride (Sigma). Sequential concentrations of ferric sulfate (Sigma) were used as an external standard [17].
Statistical analysis
The obtained information was expressed as mean±standard deviation. The statistical variance among groups was analyzed via one-way analysis of variance (ANOVA), and Tukey post hoc test was used to determine the difference between the groups. SPSS version 16 (SPSS Inc., Chicago, IL, USA) was used for data analysis and P<0.05 was considered significant.
Results
Pancreas weight
In the present study, the effective dose of nicotine (2.5 mg/kg) caused a significant decrease in the pancreas weight of the mice compared to the normal control group (P<0.05). The mean weight of pancreas was not significant in any of genistein groups compared to the normal control group (P>0.05). Moreover, pancreas weight was increased in all genistein and genistein+nicotine groups compared to the nicotine control group (P<0.05) (Fig. 2).
Serum nitrite oxide measurement
The mean of nitrite oxide in blood serum increased significantly in the nicotine control group compared to control normal group (P<0.05). The mean nitrite oxide level was not significant in any of genistein groups compared to the normal control group (P>0.05). The mean nitrite oxide level in blood serum decreased in all genistein and genistein+nicotine groups compared to the nicotine control group (P<0.05) (Fig. 3).
Serum biochemical assay
The insulin level in blood serum decreased significantly in the nicotine control group compared to the control normal group (P<0.05). The insulin in blood serum was not significant in any genistein groups compared to the normal control group (P>0.05). The insulin level in blood serum increased in all genistein and genistein+nicotine groups compared to the nicotine control group (P<0.05) (Fig. 4).
Morphometric measurements
The mean diameter and the number of pancreatic islets increase significantly in all the genistein and genistein + nicotine groups compared to the nicotine control group (P<0.05). The mean diameter and the number of pancreatic islets were not significant in any genistein groups compared to the normal control group (P>0.05). Further, nicotine caused a significant decrease in the mean diameter of pancreatic islets in the nicotine control group compared to the control normal group (P<0.05) (Fig. 5).
Histopathological observations
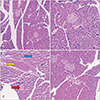
Histopathological analysis showed normal pancreas structure in the control normal and genistein and genistein+nicotine groups. After treatment with nicotine, the pancreas showed evident changes and injuries. These anomalies included a vacuolization in tissues of the pancreas, decrease in small of islet and blooding in pancreas tissue. Treatment with nicotine+genistein showed that genistein reduced pancreas injury due to the nicotine toxicity (Fig. 6).
Total antioxidant capacity
The outcomes displayed that the total antioxidant capacity serum level reduced significantly in the nicotine control group compared to the normal control group (P<0.05). Genistein increased total antioxidant capacity levels in the treated rats of complete doses compared to the nicotine control groups (P<0.05). The total antioxidant capacity level enhanced significantly in all genistein+nicotine groups compared to the nicotine control group (P<0.05) (Fig. 7).
Discussion
The nicotine content of cigarettes has important biological effects and plays a key role in the pathogenesis of many diseases. Thrower [18] have shown that nicotine is a major contributor to pancreatic anomalies. The present study evaluates the effects of genistein against toxic properties of nicotine to the pancreas weight, total antioxidant capacity, nitrite oxide, diameter, and the number of pancreatic islets and serum level of insulin. The results of the analysis of pancreas weight demonstrated a significant reduction in the pancreas weight between the normal control group and nicotine control group. In all genistein and nicotine+genistein groups, the mean pancreatic weight were increased significantly compared to the nicotine control group. Nicotine may suppress appetite mice through stimulation of melanocortin-4 receptors expressed on hypothalamic neurons [19]. Nicotine administration reduces pancreas weight by reducing starvation and enhancing thermogenesis [20]. The increase in pancreas weight might be indicative of improved nutrition of the animals under treatment with genistein [21]. The nitrite oxide serum results showed that administration of nicotine increased significantly nitrite oxide in the nicotine control group compared to the normal control group. Genistein+nicotine decreased the effects of nicotine in increasing nitrite oxide. ROS production induced by nicotine can stimulate the activity of caspase-2 and the production of nitrite oxide. In the human body, absorption of nicotine seems to be tracked through the rise of serum nitrite oxide level [22]. Genistein can perform as a phytoestrogen and antioxidant [23]. It seems that genistein suppresses nitrite oxide production in mouse macrophages, which is in line with the results of the current study [23]. Scuro et al. [24] reported that genistein could suppress the expression of nitrite oxide. The current results showed a significant decrease in insulin level in the nicotine control group compared to the normal control group. Administration of genistein+nicotine increased insulin, which shows genistein effects against the toxicity induced by nicotine impairs insulin action and prompts the body to make extra glucose. Guo et al. [25] reported that genistein administration increased insulin production in mice and showed that genistein effects on diabetes might be related to direct activities on the pancreas, confirming the results of the present study. The results of the current study can reflect the antioxidant effects of genistein. Genistein can inhibit the activities of the insulin receptor is tyrosine kinase receptors [26]. In this study, changes in pancreatic tissue due to nicotine in nicotine control group such as hyperemia in the pancreas vessels, vacuolization, and damage to pancreatic cells were observed. After genistein treatment, these symptoms were minimized, probably due to the antioxidant effects of genistein. Nicotine can cause generation of free radicals in tissues [27]. The histological outcomes in the current study showed an increase in the diameter and number of pancreatic islets in the genistein groups. Genistein can act as B cells growth factor, suggesting a new mechanism for the antidiabetic effect of this agent [28]. The results of the current study revealed that nicotine reduced the total serum antioxidant capacity level. The total antioxidant capacity level improved significantly in the genistein and genistein+morphine groups compared to the nicotine control group. The reduction at a total antioxidant capacity level in the current study shows the oxidative stress effects from nicotine at pancreases strictures. This result demonstrates the growth in the rate of ROS and a reduction in the action of antioxidant enzymes like total antioxidant capacity. The antioxidant volume of pancreases tissue by means of the high metabolism and great rate of unsaturated fatty acids in the membrane of its cells can be very damaging [23]. In the present study, improved levels of total antioxidant capacity in mice treated with genistein highlight the antioxidant properties of genistein. In general, the results of the present research showed that administration of genistein, as a potent antioxidant, to the animals in the nicotine control group could affect the performance of pancreas. Based on the obtained results, the possible antioxidant impacts of genistein have been effective in insulin. Moreover, the effects of genistein against free radicals can improve the performance and structure of pancreas against the destructive effects of nicotine.
The present study showed that genistein could improve significantly some of the pancreas damages against the toxicity effects of nicotine in mice. The antioxidant properties of genistein can be the main reason for its positive effect on pancreas parameters. However, further research is needed to describe its precise mechanism of action.






 XML Download
XML Download