

 PDF
PDF Citation
Citation Print
Print



Abbreviations
BALF
bronchoalveolar lavage fluid
F
fusion
F-DNA
respiratory syncytial virus fusion protein encoding plasmid DNA
FI-RSV
formalin-inactivated respiratory syncytial virus
F-VLP
virus-like particles presenting respiratory syncytial virus fusion protein
GC
germinal center
H&CR
hematoxylin and congo red
MLN
mediastinal draining lymph node
Naïve inf
naive mice with RSV infection
PAS
periodic acid-Schiff stain
pDC
plasmacytoid dendritic cell
RSV
respiratory syncytial virus
VED
vaccine-enhanced disease
INTRODUCTION
Human respiratory syncytial virus (RSV) causes bronchiolitis in infants and young children as well as severe respiratory illness in the elderly and immunocompromised adults (12). Human trials of formalin-inactivated RSV (FI-RSV) vaccines resulted in disastrous consequences of causing vaccine-enhanced disease (VED) during winter season (3). Mice immunized with FI-RSV in the alum formulation represented a model of recapitulating VED, suggesting that non-replicating vaccine-induced Th2 biased immune responses are associated with enhanced RSV disease (456).
DNA vaccines direct the production of mRNA and endogenously synthesize the vaccine proteins in vivo, subjecting to host post-translational modifications and making an authentic immunogen. This in vivo synthesis of RSV F proteins by DNA vaccines attributes the induction of Th1 profile immune responses of IgG2a isotype anti-F IgG and neutralizing Abs as well as cytotoxic CD8 T cells (7). RSV F-DNA boost vaccination was shown to switch preexisting Th2 anti-F responses toward Th1 type immune responses (7). Mice immunized with recombinant vaccinia virus expressing RSV attachment glycoprotein G (r-vaccinia G) were shown to develop eosinophilia similar to FI-RSV vaccination, whereas r-vaccinia F immunization of mice did not induce pulmonary eosinophils upon RSV challenge (89). Despite the difference in pulmonary eosinophil infiltration, r-vaccinia G or F immunization of mice displayed enhanced RSV disease such as weight loss after challenge (10111213). Strong induction of IFN-γ responses representing a Th1 profile was observed in mice with r-vaccinia G or F immunization (12131415). Thus, both endogenous and exogenous expression of RSV immunogens may have the potential to cause VED after RSV challenge. However, it is less well known whether RSV F-DNA vaccination would induce VED upon RSV challenge.
Virus-like particle vaccines presenting RSV F alone (F-VLP) were demonstrated to confer protection against RSV without displaying VED in mice (1617). Since gene-based in vivo immunogen synthesizing approaches are considered as an RSV vaccine platform, it is important to better understand protection and disease after RSV F-DNA vaccination. In this study, we investigated the immunogenicity, protection, VED, and histopathology in a mouse model after vaccination with F-DNA, F-VLP, or FI-RSV followed by RSV challenge. This study suggests that F-DNA vaccine is less immunogenic and protective, causing overt weight loss and pulmonary histopathology compared to F-VLP upon RSV challenge in mice. Potential immune parameters possibly correlated with RSV disease were investigated and discussed.
MATERIALS AND METHODS
Cells, virus, and RSV vaccines
The RSV A2 strain was originally gifted from Dr. Barney Graham. Hep-2 cells were obtained from American Type Culture Collection and maintained in DMEM media. The expression plasmid encoding a full-length human codon -optimized RSV A2 F within the pcDNA3 plasmid was described in previous studies (1819). F-VLP vaccine was produced in insect cells by co- expressing RSV F and influenza virus matrix 1 (M1) core protein and characterized as previously described (20). Briefly, Sf9 insect cells were co-transfected with recombinant baculoviruses expressing M1 and RSV F protein in serum-free SF900-II medium. The culture supernatants were collected by centrifugation (6,000 rpm, 20 min) to remove insect cells. The cleared supernatants containing F-VLPs were purified by ultracentrifugation. The purified F-VLPs were resuspended in PBS. F-DNA vaccine was purified using Qiagen Plasmid Mega Kit (Qiagen, Hilden, Germany). RSV was grown in HEp-2 cells, inactivated with formalin (1:4,000 vol/vol) for 3 days at 37°C, purified using ultracentrifugation to remove cell culture supernatants and cellular proteins, and finally confirmed by a modified immuno-plaque assay as previously described (20). Briefly, the infected Hep-2 cell plates were incubated at 37°C, 5% CO2 for 16 h and then viral supernatants were removed. After overlay with a mixture of 1% agarose and DMEM media, the infected Hep-2 cell plates were further incubated for 3 days, the infected Hep-2 cell plates were fixed with 10% formalin and then overlay agarose was removed. Viral plaques by RSV A2 were immune-stained with 131-2a monoclonal Ab and DAB substrate (Invitrogen, Carlsbad, CA, USA).
Vaccination, sample collection, and virus challenge
For F-VLP and F-DNA vaccine groups, BALB/c mice (6 to 8 wk old, n=5, duplicate) were intramuscularly primed and boosted with 20 μg F-VLP or 100 μg plasmid F-DNA at 0, 3 wk. Second boost vaccination was followed with a half dose of F-VLP (10 μg) or F-DNA (50 μg) respectively at wk 6. The FI-RSV control group was intramuscularly primed (wk 0) and boosted (wk 3) with FI-RSV (2 μg) in alum (50 µg) adjuvant. Each group of immunization and challenge experiments was duplicated. Naïve or vaccinated mice were challenged intranasally with RSV A2 strain (3.3×105 PFU, plaque-forming units) 20 wk after 2nd boost. The individual lungs and bronchoalveolar lavage fluid (BALF) samples were collected day 5 post-challenge to determine inflammatory cellular responses (1718). All animal experiments were approved by the Georgia State University Institutional Animal Care and Use Committees (IACUC A18001) and performed in accordance with relevant guidelines and regulations.
Assays for Ab responses, RSV neutralizing activity, and lung viral titers
RSV specific Abs (IgG, IgG1, and IgG2a) were determined by ELISA using RSV F protein antigen (BEI) and presented in concentrations based on standard IgG and isotypes as described (17). Neutralizing Ab titers in mouse sera were measured using the red fluorescent RSV A2-K-line19F (200 PFU per well) and lung viral titers by an immunoplaque assay as described (2122).
Pulmonary histology of RSV-infected mice
Individual lungs were immersed in 10% neutral buffered formalin for 24 h, embedded in paraffin, sectioned, and stained with H&E, periodic acid-Schiff stain (PAS) or hematoxylin and congo red (H&CR) as described (222324). For numerical assessment of histopathology and pneumonia in lung tissues, the bronchioles, vessels and interstitial space were initially scored on a scale of 0–3 by blinded observers with the severity scoring system as previously described (2325). A score 1 was assigned when the epithelial lines, vessels, and interstitial surrounding spaces have few infiltrating cells, a score 2 when the epithelial lines and surrounding spaces contain focal aggregates of infiltrating cells or the structure is cuffed by one definite layer of infiltrating cells, and a score 3 when structure is obviously cuffed by two or more definite layers of infiltrating cells with focal aggregates.
Cell preparations and flow cytometry
BALFs were harvested from the lung airways by infusion with PBS via trachea using a 25-gauge catheter (26). Lung tissues were homogenized, passed through a cell strainer, and spun on 44/67% Percoll gradients to collect lung cells. Cellular phenotypes were determined by flow cytometric analysis using cell surface marker Abs specific for CD3, CD4, CD8, CD11b, CD11c, CD45, F4/80, Siglec F, DX5, and Ly6c (eBioscience, San Diego, CA, USA or BD Pharmingen, San Jose, CA, USA) as previously described (172728). Cells from BALF and lungs were stimulated with the synthetic F85-93 (KYKNAVTEL) peptide (29) for CD8 T cell activation. Intracellular cytokine-producing cells stained with monoclonal IFN-γ and TNF-α and cell phenotypic marker Abs were acquired by the Becton-Dickinson LSR-II/Fortessa flow cytometer and data analyzed by Flowjo software (Tree Star Inc., Ashland, OR, USA). In the determination of cellular phenotypes, the fraction (%) of each cell phenotypes obtained by flow cytometry was multiplied by the total cell numbers counted in the cell preparations from the lung tissues and BALF.
RESULTS
F-VLP or F-DNA immunization induces IgG2a isotype dominant Ab responses
To compare the immunogenicity of exogenous F-VLP and endogenous F-DNA vaccine platforms, RSV F protein antigen-specific Abs were determined in immune sera from immunized mice (Fig. 1). Prime dose of F-DNA vaccine induced low levels of RSV F specific IgG Abs, requiring multiple immunizations. Whereas, the F-VLP group of mice induced substantial levels of IgG and IgG2a isotype Abs specific for RSV F protein after prime immunization. Significant levels of RSV F protein specific IgG and IgG2a Abs were induced after second boost with F-DNA vaccine although they were lower than those induced by F-VLP boost (Fig. 1A-C). Both F-VLP and F-DNA vaccines preferentially raised IgG2a isotype dominant Ab responses (Fig. 1B) as evidenced by high IgG2a/IgG1 ratios of Ab responses (Fig. 1D). In contrast, FI-RSV vaccination induced highest levels of IgG1 isotype Abs (Fig. 1C), suggesting Th2 immune responses. RSV neutralizing activity as measured by reduction percentages of fluorescent RSV virus infection and plaque assays was observed at higher levels in both immune sera from the F-VLP and F-DNA groups compared to naïve sera (Fig. 1E). Immune sera of alum adjuvanted FI-RSV vaccinated mice exhibited highest levels of RSV neutralizing activity (Fig. 1E).
Figure 1
F-DNA and F-VLP vaccines induce RSV F specific IgG2a dominant and neutralizing Ab responses. (A) IgG, (B) IgG2a isotype, and (C) IgG1 isotype Abs in sera. Each value represents the mean±SEM in duplicate. (D) RSV neutralizing activity of immune sera. Serially diluted mouse sera at wk 3 after 2nd boost were inactivated and used to determine RSV neutralizing activity. Statistical significances (GraphPad InStat software) are indicated in the comparison groups marked.
Naïve, unimmunized mice; FI-RSV (p), FI-RSV prime; FI-RSV (b), FI-RSV prime-boost; F-VLP (p), F-VLP prime; F-VLP (b), F-VLP prime, 1st and 2nd boost immunized mice; F-DNA (p), F-DNA prime; F-DNA (b), F-DNA prime, 1st and 2nd boost immunized mice.
**p<0.01; ***p<0.001.

F-DNA immunization causes RSV disease of weight loss after challenge
To assess the protective efficacy of F-VLP and F-DNA vaccines, immunized mice were challenged with RSV A2 at 20 wk after second boost. F-VLP immunized mice did not show body weight loss, but approximately 10% body weight loss was observed in the naïve mice infected with RSV (Naïve inf, Fig. 2A). In contrast, F-DNA immunized mice displayed significant weight loss up to 15% (Fig. 2A). The highest RSV titers in lungs were detected in unvaccinated naïve mice on day 5 post infection (Fig. 2B). Approximately 10-fold lower lung RSV titers were observed in F-DNA immune mice compared to naïve infection mice, significantly clearing lung viral loads. The F-VLP group lowered lung viral titers by over 200 folds near to the detection limit (Fig. 2B). The alum adjuvanted FI-RSV group effectively cleared lung viral loads below the detection limit (Fig. 2B).
Figure 2
F-DNA immune mice display weight loss despite control of lung viral loads after challenge. Unvaccinated naïve or vaccinated BALB/C mice were intranasally challenged with RSV A2 (3.3×105 PFU) at 20 wk after 2nd boost. (A) Changes in body weight. Differences are marked between F-VLP and F-DNA. (B) RSV titers in lungs. Lungs from individual mouse (n=10) were collected on day 5 post challenge and titers determined by an immunoplaque assay. The same vaccine groups as in the Fig. 1 were challenged with RSV. Statistical significances are indicated between naive inf and immune mice and between F-VLP and F-DNA.
LOD, limit of detection; FI-RSV, FI-RSV boost immune mice; F-VLP and F-DNA, F-VLP or F-DNA 2nd boost immunized mice on day 5 post RSV challenge.
*p<0.05; **p<0.01; ***p<0.001.

F-DNA vaccination induces infiltrating histopathology upon RSV challenge
The lung tissue sections were examined by staining with H&E, PAS, or H&CR at 5 days after RSV challenge (Figs. 3 and 4). As expected, mice that were immunized with FI-RSV showed the highest degree of pulmonary histopathology. After RSV challenge, F-DNA vaccination was found to induce significant infiltrates in the interstitial spaces of the lungs, which were similar to or slightly lower than FI-RSV (Fig. 3A and B). Moderate levels of infiltrates were observed in the blood vessels and airways from F-DNA immune mice (Fig. 3D). Meanwhile unvaccinated naïve mice exhibited moderately higher levels of infiltrating histopathology in the interstitial spaces after RSV infection (Fig. 3A and B). F-VLP immune mice did not show overt histopathology infiltrates upon RSV infection.
Figure 3
F-DNA immunization causes pulmonary infiltrates and pathology after challenge. Lung tissues for histology (scale bars, 100 µm) were collected on day 5 post challenge. (A) Microscope pictures of H&E stained tissue sections. (B-D) Inflammation scores on a scale of 0 to 3 as diagnostic criteria (n=5). (B) Interstitial spaces. (C) Airways. (D) Blood vessels. Groups are the same as described in Fig. 2. Statistical significances are marked by lines among the groups for comparison.
*p<0.05; **p<0.01; ***p<0.001.
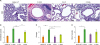
Figure 4
F-DNA immune mice display moderate eosinophilia after challenge. (A) Mucus positive PAS staining (scale bars, 100 µm). An arrow marks dense PAS staining area. (B) Eosinophilic H&CR staining (scale bars, 20 µm). The inserts in H&CR images are details of arrow areas with eosinophilic stains in lungs. (C) Percentages for bronchiolar PAS positive mucus. Each symbol in mucus production percentage represents one airway of 10 individual airways. (D) Eosinophils. The degrees of eosinophilia were expressed as numbers of eosinophils (H&CR stains) present per 400× field. Bars indicate significant differences between groups. Groups are the same as described in the Fig. 2.
*p<0.05; **p<0.01; ***p<0.001.
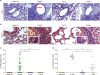
Mucus production and eosinophilia are also considered RSV disease parameters. FI-RSV immunization showed the highest levels of PAS positive mucus production and H&CR positive eosinophils upon RSV challenge (Fig. 4). F-DNA immunization caused low levels of eosinophils (Fig. 4B and D). The naïve and F-VLP groups did not display PAS positive spots and eosinophils in the lung sections (Fig. 4A and C)
Inflammatory innate immune cells are recruited into the lungs in F-DNA immune mice upon RSV challenge
Cellular phenotypes in lung (Fig. 5) and BALF (Fig. 6) were analyzed by flow cytometry using cell type-specific marker Abs at 5 days after RSV challenge. Alum-adjuvanted FI-RSV immune mice showed the highest levels of eosinophils (CD11b+CD11c−SiglecF+) in lung and BALF samples as well as plasmacytoid dendritic cells (pDC, B220+CD11c+F4/80−) in BALF. F-DNA immunized mice displayed higher levels of NK cells (CD3−DX5+), monocytes (CD11b+F4/80+Ly6Chigh), eosinophils, and pDC in the lungs compared to F-VLP or naïve infection mice. Particularly, eosinophil and monocyte populations were increased by approximately 3-fold in F-DNA mice than those in F-VLP mice. We also observed increased levels in monocytes and pDCs in airway BALF samples from F-DNA immunized mice, compared to those in F VLP (Fig. 6).
Figure 5
F-DNA immunization induces inflammatory innate and CD8 T cells in lungs after challenge. Flow cytometry of immune cells was applied to numerate cellular phenotypes in lungs and MLN at 5 days after RSV challenge (n=5). (A) NK (CD3−DX5+) cells, (B) monocytes (CD11b+F4/80+Ly6Chigh), (C) eosinophils (CD11b+CD11c−SiglecF+), (D) pDC (B220+CD11c+F4/80−). (E, F) CD8 T cells of intracellular cytokine staining after stimulation of lung cells with RSV F85-93 peptide. (E) Lung IFN-γ CD8 T cells, (D) Lung TNF-α CD8 T cells. (G) GC phenotypic B cells (IgD−B220+GL7+) in MLN. Lines indicate differences between groups.
*p<0.05; ***p<0.001.

Figure 6
F-DNA immunization induces inflammatory innate cells and CD8 T cells in BALF after challenge. Flow cytometry of immune cells was applied to numerate cellular phenotypes in BALF at 5 days after RSV challenge (n=5). (A) NK (CD3−DX5+) cells, (B) monocytes (CD11b+F4/80+Ly6Chigh), (C) eosinophils (CD11b+CD11c−SiglecF+), (D) pDC (B220+CD11c+F4/80−). (E, F) CD8 T cells of intracellular cytokine staining after stimulation of BALF cells with RSV F85-93 peptide. (E) BALF IFN-γ+ CD8 T cells, (F) BALF TNF-α+ CD T cells. Lines indicate differences between groups.
*p<0.05; **p<0.01; ***p<0.001.

F-DNA is effective in inducing Th1 CD8 T cells whereas F-VLP in activating B cells upon RSV challenge
We determined the levels of CD8 T cells producing IFN-γ and/or TNF-α cytokines in the lungs at 5 days after RSV challenge by a flow cytometry assay of intracellular cytokine staining. Lung cells from F-DNA immune mice showed significantly higher levels of RSV F85-93 specific IFN-γ or TNF-α positive CD8 T cells than F-VLP, FI-RSV immune or naïve infection mice (Fig. 5E and F).
Germinal center (GC) phenotypic B cells (IgD−B220+GL7+) in the mediastinal draining lymph node (MLN) were also analyzed by flow cytometry day 5 post challenge. F-VLP immunized mice showed significantly higher levels of GC phenotypic B cells in line with increased levels of serum IgG Abs compared to F-DNA immunization. The F VLP group showed certain levels of IgA Abs in BALF but not the F-DNA group, but overall the IgA Ab levels were very low and no significant levels of IgM Ab levels were detected (data not shown).
DISCUSSION
Since the failure of alum-adjuvanted FI-RSV vaccine, non-replicating RSV subunit vaccine platforms would have potential safety concerns of causing VED in young naïve children. Here, we compared the immunogenicity, protection, and immunopathology of a full-length F-DNA vaccine for in vivo endogenous expression or F-VLP as an exogenous RSV F protein. Both RSV F-VLP and DNA vaccines were capable of inducing IgG2a isotype and RSV neutralizing Abs and controlling lung viral loads upon RSV challenge better than unvaccinated naïve mice with RSV infection. F-VLP was more effective in controlling RSV loads than F-DNA. Unexpectedly, the mice with F-DNA vaccination showed substantial weight loss and pulmonary histopathology compared to F-VLP immune or naïve mice after RSV infection. The results in this study suggest that RSV immunity in mice induced by endogenous expression of RSV F does not necessarily avoid VED after RSV infection.
The nature of endogenous synthesis of the vaccine immunogen by DNA vaccines engenders the induction of relatively weak humoral and strong cellular immune responses. RSV F specific Abs were not detected after prime with F-DNA probably due to low transgene transduction and thus multiple F-DNA immunizations required to induce substantial levels of antigen specific IgG Abs. Despite relatively low levels of IgG Abs, significant reductions of lung RSV titers were reported with soluble secreted F protein-encoding DNA vaccination being 10-fold more effective than transmembrane F protein-encoding plasmid (7). Low efficacy of lung RSV clearance in the F-DNA group might be due to the expression of membrane-anchored form. However, pulmonary inflammation was observed in animal models with F-DNA (730), FI-RSV (7), and soluble F protein (31) vaccines despite significant clearance of lung RSV loads or below the detection limit.
Different platforms of RSV vaccines influence the cellular phenotypes infiltrating into the lungs upon RSV challenge. FI-RSV or r-vaccinia G-immunized mice were shown to exhibit severe eosinophilia upon RSV challenge (2932). Substantial RSV histopathology was previously reported in mice with RSV F DNA vaccination despite the induction of neutralizing Abs, Th1 CD8 T cell responses, and reduction of RSV lung viral loads (730), consistent with this current study. Particularly in a recent study by Ma et al. (30), the groups of mice were immunized 4 times with 30 µg of F DNA vaccines containing 5 or 20 copies of built-in CpG immune stimulating motifs. Ma et al. (30) demonstrated that the pVAX1-F (5CpG motifs) group displayed a similar pattern of weight loss (>10%) as the PBS control after RSV challenge, whereas the pVAX1-F (20CpG motifs) group showed a moderate level of weight loss (<9%). In this study, we found that the F-DNA (no CpG motif) group and FI-RSV control group exhibited a substantial weight loss (>10%), which is similar to the PBS and pVAX1-F (5CpG motifs) group in the Ma et al.'s study (30). It is speculated that the overall outcomes in body weight changes after RSV challenge are similar but subtle differences might be due to the nature of F DNA vectors, doses, and immunization regimen between the two studies.
RSV F or G DNA vaccination was previously reported to infiltrate pulmonary eosinophils, NK cells, and neutrophils but histopathology was not investigated (33). F-DNA vaccine exhibited slightly higher levels of eosinophils than F-VLP vaccine after RSV challenge. However, the levels of eosinophils in the F-DNA group were significantly lower than those in FI-RSV immune mice after RSV challenge. F-DNA immune mice showed a Th1 pattern of immune responses (high IFN-γ, low IL-4, Supplementary Fig. 1), consistent with a pattern reported by Ma et al. (30). High eosinophils in the FI-RSV group were consistent with Th2 cytokine levels. Monocytes from the bone marrow traffic to peripheral tissues via the bloodstream in response to a signal of inflammation and play an important role in clearing pathogens (34). Additionally, extensive monocytes can contribute to immunopathology and degenerative disease (34). The pDCs are known to be important for producing type 1 IFNs and enhancing CD8 T cell responses (3536). Other studies reported that high levels of pDCs are involved in severe RSV disease and elevated mortality during influenza virus infection (3738). Levels of pDCs infiltrating into the lungs were shown to be correlated with FI-RSV VED (39). NK cells enhance CD8 T cells expressing IFN-γ and NK cell depletion was shown to attenuate weight loss during RSV infection (4041). Consistent with previous studies, this study suggests that recruiting NK cells, monocytes, pDCs, and eosinophils at high levels in F-DNA immune mice contribute to pulmonary histopathology during RSV infection.
There was no significant difference in the levels of IL-4 cytokines in lung cells in response to stimulation with CD4 T and CD8 T cell epitopes between the F-DNA and F-VLP groups (Supplementary Fig. 1). Unbalanced high levels of CD8 T cell responses could induce significant immunopathology. Vaccination of mice with r-vaccinia G and M2, or M282-90 peptide in adjuvants inducing strong CD8 T cell responses was shown to cause severe weight loss after RSV challenge (294243). Excess induction of IFN-γ+ T cell responses was shown to cause immune-mediated pathology in response to RSV infection (13434445). Also, high levels of TNF-α+ CD8 T cells were indicated to be an indicator for severity of pulmonary disease such as weight loss (4647). In line with previous findings, F-DNA immune mice induced high levels of RSV F specific IFN-γ+ and TNF-α+ CD8 T cells in the lungs, compared to F-VLP, FI-RSV immune or naïve mice upon RSV challenge, suggesting a potential correlation with RSV disease. Interestingly, our previous studies reported that inclusion of F-DNA in the RSV F+G VLP vaccination was effective in controlling lung viral titers and preventing pulmonary histopathology (182224). An appropriate balance of inducing CD8 T cells together with RSV neutralizing Abs would be important for conferring desirable protection avoiding VED during RSV infection.
 XML Download
XML Download