

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Tumor protein p53-regulated apoptosis-inducing protein 1 (TP53AIP1) is critical for tumor suppressor p53-dependent apoptosis [1]. TP53AIP1 is located in the mitochondrial membrane and can activate the release of cytochrome c and induce apoptosis [2]. The Ser46-phosphorylation of p53 triggers the activation of TP53AIP1, which in turn induces apoptosis in response to severe DNA damage [3]. However, TP53AIP1 is a susceptible gene and has been found to be mutated in prostate cancer [1]. The overexpression of TP53AIP1 up-regulates p53 levels in liver hepatocellular carcinoma cells, thus inducing apoptosis and cell cycle arrest [4]. It has been found that patients with non-small cell lung cancer with a lower level of TP53AIP1 have lower overall survival (OS) [5]. However, the role of TP53AIP1 in breast cancer has not yet been completely studied.
Breast cancer is a malignant tumor that originates in the ductal epithelium of the breast or breast lobule [6]. Breast cancer is one of the most frequently occurring malignant tumors in women worldwide [7]. According to the statistics from the World Health Organization (WHO), in 2000, more than 1 million new cases of breast cancer were diagnosed worldwide [8]. The standardized incidence rate was 35.66 per 100,000 people, and the standardized mortality was 12.51 per 100,000 people [8]. According to latest statistics from WHO, by 2008, there were around 1.38 million new cases of breast cancer which accounted for 23% of all new cancer cases, and breast cancer had the second highest incidence among other kinds of cancers [9]. Breast cancer is the fifth leading cause of all cancer deaths and is the most common cause of cancer related death in women [9]. In recent years, great progress has been made in the diagnosis and treatment of breast cancer, especially in the surgical treatment of breast cancer [10]. Due to deeper understanding of the biological mechanism of breast cancer, molecular targeted therapy for breast cancer is gradually becoming possible [11]. Since the prognosis of breast cancer is poor, the current treatment methods may not be effective enough to fully treat the disease. Therefore, exploring new targeted sites for treatment are of important clinical significance.
Hence, given the critical effect of TP53AP1 on tumor suppressor p53-dependent apoptosis, we investigated the molecular mechanism of TP53AP1 in breast cancer cells. This study will provide novel evidence for the diagnosis, treatment, and prognosis of breast cancer.
METHODS
The Cancer Genome Atlas (TCGA) analysis
TCGA online database (https://cancergenome.nih.gov/) was used to determine the correlation between TP53AIP1 expression and OS in patients with breast cancer. The 3-year OS was analyzed using Kaplan-Meier OS analysis.
Patients
Our study sample included 38 female patients with invasive breast cancer between the ages of 41 and 68, treated at the Third Affiliated Hospital of Chongqing Medical University (CQMU) Hospital (Chongqing, China) in 2017. No patient had undergone systemic therapy before receiving surgery. The TNM staging system was adopted. The research was approved by the Ethics Committee of the Third Affiliated Hospital of CQMU Hospital (CQMU201612017R). All patients signed the informed consent. Cancer tissues and adjacent normal tissues were obtained from each of the patients. Real-time-quantitative polymerase chain reaction (RT-qPCR), western blot, and immunohistochemical staining were performed to detect the expression levels of TP53AIP1.
Immunohistochemistry (IHC)
IHC assay was used to measure the expression of TP53AIP1 in breast cancer tissues. Tissues were immobilized with cold 4% paraformaldehyde (Sigma, St. Louis, USA), probed with rabbit anti-TP53AIP1 primary antibody (cat No. ab196765, 1:100; Abcam, Cambridge, USA) overnight at 4°C and incubated with horseradish peroxidase-conjugated goat-anti-rabbit immunoglobulin G (secondary antibody) (cat No. ab6721, 1:1,000; Abcam) for 30 minutes. Subsequently, the samples were dyed with coloring agent diaminobenzidine (Sigma) and hematoxylin. Finally, the images were observed using DMi8 optical microscope (Leica, Wetzlar, Germany) under 200-fold magnification.
Cells and treatment
MDA-MB-415 and MDA-MB-468 cells, 2 commonly used human breast cancer cell lines, were purchased from American Type Culture Collection (ATCC, Manassas, USA) and cultured at 37°C with 5% CO2. Dulbecco's modified Eagle's medium (Hyclone™; Thermo Fisher Scientific, Waltham, USA) was used as culture medium, and was supplemented with 10% fetal bovine serum (FBS), 100 U/mL streptomycin and penicillin. FBS, streptomycin and penicillin were all purchased from Biological Industries (Kibbutz Beit-Haemek, Israel). The recombined plasmid pcDNA3.1-TP53AIP1 (TP53AIP1 group) (GenePharma, Shanghai, China) and the empty vector pcDNA3.1 (negative control [NC]-vector group) (GenePharma) were transfected into MDA-MB-415 and MDA-MB-468 cells respectively using Lipofectamine 2000 (Invitrogen, Carlsbad, USA) as a transfection reagent. Control cells were those without any treatment. The transfection efficiency at different TP53AIP1 levels was detected using RT-qPCR and western blot.
Cell viability assay
Cell Counting Kit-8 (CCK-8; MedChemExpress, Monmouth Junction, USA) assay was used to determine the effect of TP53AIP1 overexpression on the viability of breast cancer cells (MDA-MB-415 and MDA-MB-468). Specifically, cells from control, NC-vector and TP53AIP1 groups were seeded into 96-well plates (2 × 103 cells/well) and cultured at 37°C for 12, 24, and 48 hours respectively. Then, 20 μL CCK-8 reagent was added to the different cells and incubated for 1 hour. Finally, the optical density values were recorded at 450 nm using a microplate reader (Biotek, Winooski, USA).
Cell cycle detection
Propidium iodide (PI; Invitrogen) staining was used to evaluate cell cycle in control, NC-vector, and TP53AIP1 groups. The cells were fixed in icy 70% ethanol overnight at 4°C, and incubated with 50 μg/mL PI and 100 μg/mL ribonuclease for 30 minutes at 37°C. The results were immediately analyzed using a fluorescence-activated cell sorting flow cytometer (BD, San Diego, USA). The percentage of cells in G0/G1, S and G2/M phases were assessed.
Apoptosis detection
Annexin V/PI assay (Biovision, Milpitas, USA) was used to analyze cell apoptosis rates in control, NC-vector, and TP53AIP1 groups. Specifically, cells were incubated with 5 μL Annexin-V and 5 μL PI for 5 minutes in the dark at 37°C and analyzed using a flow cytometer (BD).
RT-qPCR
The messenger RNA (mRNA) levels of TP53AIP1, caspase-9, caspase-3, B cell lymphoma/leukemia-2 (Bcl-2), Bcl-2-associated X protein (Bax), Ki67, and p53 in breast cancer cells (MDA-MB-415 and MDA-MB-468) and in the cells transfected with either TP53AIP1 or NC-vector were analyzed. The primers, synthesized by SBS Genetech Co., Ltd. (Beijing, China), are listed in Table 1. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as an internal control. Total RNA was extracted using Trizol reagent (Invitrogen), and then reverse transcribed to complementary DNA using Transcriptase (Roche, Kansas City, USA). Then, the PCR amplification process was conducted using LightCycler® Multiplex Masters (Roche) with LightCycler® 480II System (Roche) under the following conditions: 36 cycles at 95°C for 5 minutes (denaturation at 95°C, 34 seconds, annealing at 60°C, 34 seconds, extension at 72°C, 34 seconds), and at 72°C for 10 minutes. The results were calculated using the 2-ΔΔCq method.
Table 1
Primers used in the real-time-quantitative polymerase chain reaction assay

GAPDH = glyceraldehyde 3-phosphate dehydrogenase; TP53AIP1 = tumor protein p53-regulated apoptosis-inducing protein 1; Bax = Bcl-2-associated X protein; Bcl-2 = B cell lymphoma/leukemia-2.
![]()
Western blot
The protein levels of TP53AIP1, Bcl-2, Bax, caspase-9, caspase-3, Ki67, p53, protein kinase B (Akt), phosphorylated-Akt (p-Akt), phosphoinositide 3-kinases (PI3K), phosphorylated-PI3K (p-PI3K), and mouse double minute 2 homolog 2 in breast cancer cells (MDA-MB-415 and MDA-MB-468) and in cells transfected with either TP53AIP1 or NC-vector were measured . The cells were lysed on ice for 30 minutes using RIPA lysis buffer (Boster, Wuhan, China). The proteins were then quantified using BCA (Pierce, Boston, USA), and the protein was separated into 10 μg units using 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membranes (Thermo Fisher Scientific). Next, the membranes were blocked with 5% non-fat dry milk at 37°C for 1 hour, and reacted first, with specific primary antibodies (Abcam) overnight at 4°C and then with secondary antibodies conjugated with horseradish peroxidase (7074, 1:5,000; CST, Danvers, USA) at 37°C for 1 hour. The proteins were detected using enhanced chemiluminescence detection reagents (Pierce) and analyzed using Bio-Rad ChemiDoc XRS densitometry. GAPDH was used as a loading control.
RESULTS
TP53AIP1 is down-regulated in patients with breast cancer
At the beginning of the research, we conducted a bioinformatics analysis to determine TP53AIP1 levels using the downloaded TCGA database. The analysis showed that patients with low TP53AIP1 levels had lower OS rates, compared to those with high TP53AIP1 levels (p < 0.01, Figure 1A). To further confirm the results, we studied 38 patients with breast cancer and determined the mRNA and protein levels of TP53AIP1 in their cancer tissues and adjacent normal tissues. The overall mRNA levels of TP53AIP1 were significantly down-regulated in cancer tissues, compared with adjacent normal tissues (p < 0.050, Figure 1B). In addition, western blot assay showed that the protein levels of TP53AIP1 were down-regulated at varying degrees, and the most representative ones are shown in Figure 1C. IHC assay also demonstrated decreased TP53AIP1 in breast cancer tissues under 100- and 200-fold magnification (Figure 1D).
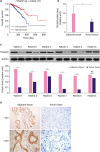 | Figure 1TP53AIP1 is down-regulated in patients with breast cancer. (A) Data from The Cancer Genome Atlas database were collected, and the data showed that breast cancer patients with low level of TP53AIP1 had lower survival rates. (B) Quantitative real-time polymerase chain reaction assay verified down-regulated overall level of TP53AIP1 mRNA in breast cancer patients. (C) Western blot showed down-regulated protein levels of TP53AIP1 in most breast cancer patients, and the representative ones are displayed. (D) Immunohistochemistry assay showed negative protein expression of TP53AIP1 in invasive breast cancer tissues and positive brown TP53AIP1 protein expression in adjacent tissues under 100- and 200-fold magnifications.TP53AIP1 = tumor protein p53-regulated apoptosis-inducing protein 1; GAPDH = glyceraldehyde 3-phosphate dehydrogenase; mRNA = messenger RNA; NS = not significant.
*p < 0.05 and †p < 0.01 vs. adjacent tissue.
|
Overexpression of TP53AIP1 inhibits viability of breast cancer cells
After transfecting either TP53AIP1 or NC-vector into breast cancer MDA-MB-415 and MDA-MB-468 cells, the transfection efficiencies were measured by determining TP53AIP1 levels. RT-qPCR and western blot assays showed that the mRNA and protein levels of TP53AIP1 were all significantly up-regulated in TP53AIP1 (MDA-MB-415 and MDA-MB-468) group, compared with the NC-vector group (p < 0.01, Figure 2A-D). CCK-8 assay showed that viabilities of TP53AIP1 (MDA-MB-415 and MDA-MB-468 cells) were markedly inhibited at 24 and 48 hours (Figure 2E and F).
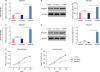 | Figure 2Overexpression of TP53AIP1 inhibits viability of breast cancer cells. RT-qPCR (A) and Western blot (B) were used to show increased mRNA and protein levels of TP53AIP1 in TP53AIP1 (MDA-MB-415) group. RT-qPCR (C) and western blot (D) were used to show increased mRNA and protein levels of TP53AIP1 in TP53AIP1 (MDA-MB-468) group. Cell Counting Kit-8 assay was used to show inhibited cell viabilities in TP53AIP1 (MDA-MB-415) group (E) and TP53AIP1 (MDA-MB-468) group (F).TP53AIP1 = tumor protein p53-regulated apoptosis-inducing protein 1; GAPDH = glyceraldehyde 3-phosphate dehydrogenase; mRNA = messenger RNA; RT-qPCR = real-time-quantitative polymerase chain reaction; NS = not significant; OD = optical density; NC = negative control.
*p < 0.01 vs. NC-vector group.
|
Overexpression of TP53AIP1 promotes cell cycle arrest and apoptosis of breast cancer cells
Flow cytometry was used to show percentages of cells in different cell cycle phases and apoptotic cells rates using PI cell cycle assay and Annexin V/PI apoptosis assay. The results demonstrated that cell cycle arrest occurred between G0/G1 phase and S phase in TP53AIP1 (MDA-MB-415) group, with more cells staying in the G0/G1 phase and less cells transferring into S phase, compared with each NC-vector group (Figure 3A). The apoptosis rates were drastically elevated in TP53AIP1 (MDA-MB-415) group, compared with each NC-vector group (Figure 3B). In MDA-MB-468 cells TP53AIP1 had similar results on cell cycle arrest (Figure 3C) and apoptosis (Figure 3D).
 | Figure 3Overexpression of TP53AIP1 promotes cell cycle arrest and apoptosis of breast cancer cells. Cell cycle arrest (A) and apoptosis rates (B) were found to be promoted in TP53AIP1 (MDA-MB-415) group. Cell cycle arrest (C) and apoptosis rates (D) were found promoted in TP53AIP1 (MDA-MB-415) group and (MDA-MB-468) group. The rates were determined using flow cytometry.TP53AIP1 = tumor protein p53-regulated apoptosis-inducing protein 1; NC = negative control; FL2 = fidgetin-like 2; NS = not significant.
*p < 0.01 vs. NC-vector group.
|
Overexpression of TP53AIP1 promotes cell cycle arrest and apoptosis of breast cancer cells by regulating cell cycle and apoptosis related factors
Since overexpression of TP53AIP1 significantly promoted cell cycle arrest and apoptosis of breast cancer cells, we also studied their molecular mechanisms by detecting changes in the expression of cell cycle and apoptosis-related factors due to TP53AIP1 overexpression. RT-qPCR and western blot assays demonstrated that both the mRNA and protein levels of Bax and p53 were significantly increased, while Bcl-2 and Ki67 levels were significantly decreased in TP53AIP1 (MDA-MB-415 and MDA-MB-468) group, compared with each NC-vector group (p < 0.01, Figure 4). In addition, protein levels of cleaved-caspase-9 and cleaved-caspase-3 were noticeably increased in the TP53AIP1 (MDA-MB-415 and MDA-MB-468) group (p < 0.01, Figure 4E and J).
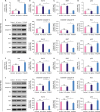 | Figure 4Overexpression of TP53AIP1 promotes cell cycle arrest and apoptosis of breast cancer cells by regulating cell cycle and apoptosis related factors. The mRNA levels of Bax and p53 increased, and Bcl-2 and Ki67 levels decreased in TP53AIP1 (MDA-MB-415) (A-D) and TP53AIP1 (MDA-MB-468) (F-I) groups. The protein levels of cleaved-caspase-3, cleaved-caspase-9, Bax and p53 increased and Bcl-2 and Ki67 levels decreased in TP53AIP1 (MDA-MB-415) (E) and TP53AIP1 (MDA-MB-468) (J) groups.TP53AIP1 = tumor protein p53-regulated apoptosis-inducing protein 1; GAPDH = glyceraldehyde 3-phosphate dehydrogenase; Bcl-2 = B cell lymphoma/leukemia-2; Bax = Bcl-2-associated X protein; mRNA = messenger RNA; NC = negative control; NS = not significant.
*p < 0.05 and †p < 0.01 vs. NC-vector group.
|
Overexpression of TP53AIP1 promotes cell cycle arrest and apoptosis of breast cancer cells via inhibiting the PI3K/Akt pathway
Since PI3K/Akt pathway is an important signaling pathway in cell growth, we studied the effect of TP53AIP1 overexpression on the PI3K/Akt pathway. The western blot results showed that p-PI3K/PI3K ratio, p-Akt/Akt ratio and mouse double minute 2 homolog (Mdm2) levels were all effectively suppressed in TP53AIP1 (MDA-MB-415 and MDA-MB-468) groups, compared with each NC-vector group (p < 0.01, Figure 5).
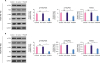 | Figure 5Overexpression of TP53AIP1 promotes cell cycle arrest and apoptosis of breast cancer cells via inhibiting the PI3K/Akt pathway. The percentage of p-PI3K/PI3K, p-Akt/Akt, and Mdm2 levels were suppressed in TP53AIP1 (MDA-MB-415) (A) and TP53AIP1 (MDA-MB-468) (B) groups.TP53AIP1 = tumor protein p53-regulated apoptosis-inducing protein 1; GAPDH = glyceraldehyde 3-phosphate dehydrogenase; NC = negative control; NS = not significant; PI3K = phosphoinositide 3-kinases; Akt = protein kinase B; p-Akt = phosphorylated-Akt; p-PI3K = phosphorylated-PI3K; Mdm2 = mouse double minute 2 homolog.
*p < 0.01 vs. NC-vector group.
|
DISCUSSION
As one of the most frequently occurring malignant tumors in women worldwide [12], breast cancer is a serious threat to the health of women [13]. Through bioinformatics analyses and research on clinical breast cancer tissues, we found that TP53AIP1 was significantly decreased in breast cancer tissues. Breast cancer patients with low TP53AIP1 levels had lower survival rates than those with higher levels of TP53AIP1. This difference suggests that TP53AIP1 may be a tumor suppressor gene in breast cancer and that its role in breast cancer is similar to that in several other tumors [4,5]. In this study, transgenic technology was used to enhance the expression of TP53AIP1 in breast cancer cells lines, MDA-MB-415 and MDA-MB-468. We found that TP53AIP1 overexpression inhibited viability of breast cancer cells, blocked the cell cycle, and promoted cell apoptosis, indicating that the mechanism, through which TP53AIP1 inhibits the proliferation of breast cancer cells may be related to cell cycle arrest and apoptosis promotion.
Cell cycle refers to all the processes from the end of one cell division to the end of next cell division [14]. It contains 2 phases, interphase and division phase [15]. A classic cell cycle includes G1, S, G2, and M phases [16]. Cell cycle regulation, mainly achieved through the retention of G1 phase, is of great significance in the understanding of growth and development of organisms and controlling tumor growth [17]. Finding ways to promote tumor cell stagnation is critical in controlling tumorigenesis. In our study, overexpression of TP53AIP1 induced cell cycle arrest in G1 phase in breast cancer cells, indicating that TP53AIP1 is a potential factor in the inhibition of tumor growth.
Apoptosis, also known as programmed cell death, is a special type of cell death in the biological world [18]. Apoptosis activates the membrane signaling system and apoptotic procedural genes, and when it is induced by certain physiological or pathological factors, apoptosis ultimately leads to self-destruction and death of cells via certain processes [19]. Studies have shown that apoptosis is regulated by a series of genes can be divided into 2 groups based on their effects—apoptosis inhibitory factors and apoptosis promoting factors. Bcl-2 and Ki67 are apoptosis inhibitory factors. Bax and p53 are apoptosis promoting factors [20]. Bax was the first discovered pro-apoptotic factor in the Bcl-2 family [21]. In p53-dependent apoptosis, p53 can specifically inhibit Bcl-2 expression and promote Bax expression, thus eventually promoting apoptosis [22]. Ki67, found to be positively expressed in breast cancer, is a nuclear protein associated with proliferating cells [23]. TP53AIP1 is the apoptosis inducing protein that is directly regulated by p53. Previous research showed that overexpression of Bcl-2 blocks the proapoptotic activity of TP53AIP1 [24]. In this study, we discovered that the over-expression of TP53AIP1 significantly suppressed expression levels of Bcl-2 and Ki67 and promoted the expression of Bax and p53, thus activating apoptosis progression in breast cancer cells. The caspase family is structurally related to cysteine protease in the cytoplasm. The important common feature of caspase family members is the ability to specifically cleave peptide bonds after aspartic acid residues. caspase-3 and caspase-9 are the most important effectors in the caspase family. Caspase-9 contributes to the initiation of apoptosis, while caspase-3 contributes to the execution of apoptosis. Activated caspase-9 could further activate the downstream target, caspase-3, to initiate the apoptosis program. In this study, the activation of caspase-9 and caspase-3 were both consistently facilitated by TP53AIP1 over-expression in breast cancer cells. Therefore, the mechanism of TP53AIP1 inducing cell cycle arrest and apoptosis may be dependent on the regulation of apoptosis-related factors, such as p53, Bax, Bcl-2, and Ki67.
The PI3K/Akt pathway is a critical signal transduction pathway that plays a role in tumorigenesis [25]. Studies have shown that the PI3K/Akt pathway is involved in the apoptosis regulated by caspase-9 activation in breast cancer, colon cancer and prostate cancer cells [26,27]. Akt can be phosphorylated by PI3K and activated Akt either activates or inhibits downstream target proteins through phosphorylation, which in turn regulate cell proliferation, differentiation, apoptosis and migration [28]. Activation of Akt can dissociate the Akt-Mdm2 complex and the isolated Mdm2 enters the nucleus and forms a p53-Mdm2 complex with p53, thus inhibiting the transcriptional activity of p53 and eventually leading to the accelerated apoptosis of the tumor cells [29,30]. During our research, we discovered that the over-expression of TP53AIP1 significantly reduced the phosphorylation of PI3K and Akt as well as the expression of Mdm2 in breast cancer MDA-MB-415 and MDA-MB-468 cells. Hence, the mechanism of TP53AIP1 inhibiting the proliferation and growth of breast cancer cells may be related to cell cycle arrest, apoptosis promotion and altering the PI3K/Akt pathway.
There were some limitations in our study; for example, the follow-up period of the cases used for the survival analysis was short. Thus, further investigation is required.
In conclusion, TP53AIP1 acts as a tumor suppressor, and its overexpression inhibits cell proliferation and cell cycle progression and promotes apoptosis in breast cancer cells via regulating the PI3K/Akt pathway. Our results may provide a theoretical basis for the molecular targeted therapy of breast cancer.
 XML Download
XML Download