

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Bones are an organ that changes over time during the process of production, growth, and absorption over a lifetime. Approximately 10% of an adult bone is replaced with new bone each year.[1] The bone density is highest in people in their 20s to 30s and decreases gradually after that. In women, the bone mineral density decreases rapidly during the first 5 years of menopause.[2] Excessive bone loss is a major cause of osteoporosis, which is becoming a public health problem because of the increased frequency of osteoporotic fractures in elderly people.[3] Osteoporosis is an age-related disorder that is characterized by bone loss and deterioration of the bone structure, particularly affecting postmenopausal women.[4]
Bone homeostasis/remodeling is important for maintaining the bone mass and quality. This process requires a balanced action of bone-resorbing osteoclasts and bone-forming osteoblasts.[5] Typical bone diseases, such as osteoporosis, Paget's disease, and rheumatoid arthritis, have the symptoms of decreased bone mass because of the increased number and activity of osteoclasts.[6] Osteoclasts are large multinucleated cells (MNCs) that remove the old/weakened bones by acid decalcification and proteolytic degradation.[7] They are differentiated from the monocyte-macrophage of a hematopoietic lineage. Osteoclast formation is controlled by 2 cytokines, macrophage colony-stimulating factor (M-CSF) and receptor activator of nuclear factor-κB ligand (RANKL), which are secreted by osteoblasts/activated T cells.[8] M-CSF acts on osteoclast precursor cells to induce signaling related to cell survival and promotes the expression of RANKL receptor, RANK.[9] RANKL is also known as the tumor necrosis factor-related activation-induced cytokine or osteoclast-differentiation factor. Binding of RANKL and RANK ultimately leads to the formation of osteoclasts by increasing the expression of nuclear factor of activated T cells 1 (NFATc1), a key transcription factor for osteoclastogenesis.[10]
Lichen is a symbiotic relationship between fungi (mycobiont) and photosynthetic organisms (photobionts) that use each other's metabolites to generate a range of secondary metabolites.[1112] Their metabolites have been discovered over the past decade and are being assessed to find new bioactive compounds.[1314] Recently, various species of lichen were collected and bioactive extracts were obtained. In addition, the extracts of Flavoparmelia sp. (EFV) inhibited osteoclast formation. This study investigated the mechanisms by which EFV inhibits osteoclast formation.
METHODS
1. Preparation of Flavoparmelia sp. extracts
Flavoparmelia sp. was collected from Peru in May of 2017 during a field trip of San Tuario, Muruhuay, Acobamba, Junin organized by Prof. Rebeca Magdalena Pavlich Herrera at Peruvian University Cayetano Heredia, Peru. The field trip was conducted in the frame of an internal joint program between Korea and Peru, supported by Korea National Research Foundation. The field studies did not involve endangered or protected species. Duplicates were deposited at the Korean Lichen and Allied Bioresource Center in the Korean Lichen Research Institute, Sunchon National University (SNCU), Korea. The air-dried Flavoparmelia sp. (10 g) were extracted twice with 2 L methanol at room temperature for 48 hr using sonication. The extract was filtered then concentrated under vacuum at 40℃ using a rotary evaporator. The extract was subjected to high performance liquid chromatography (HPLC) analyses (LC-20A; Shimadzu, Kyoto, Japan) on a YMC-Pack™ ODS-A (150×3.9 mm I.D.; YMC, Kyoto, Japan) reverse-phase column containing fully end-capped C18 material (particle size, 5 µm; pore size, 12 nm). Elution was performed at a flow rate of 1 mL/min under the following conditions before subsequent injection: column temperature, 40℃; and solvent system, methanol: water:phosphoric acid (80:20:1, v/v/v). The analyses were monitored using a photodiode array detector (SPD-M20A; Shimadzu) over the range, 190 to 800 nm, throughout the HPLC run. The observed peaks were scanned between 190 and 400 nm.
2. Cell culture and osteoclast differentiation
This study was conducted in strict accordance with the recommendations contained in the Standard Protocol for Animal Study of SCNU. The protocol was approved by the SCNU Institutional Animal Care and Use Committee (IACUC) with Permit No. SCNU IACUC 2016-06. All efforts were made to minimize suffering.
All cells were cultured in a 5% CO2 at 37℃. The culture medium was replaced with fresh medium every 3 days. Bone marrow cells (BMCs) were isolated from the femurs and tibias of 5-week-old male ICR mice (n=2; RaonBio Inc., Yongin, Korea). The BMCs were incubated with 10 ng/mL M-CSF (PeproTech, Rocky Hill, NJ, USA) for 16 hr in α-minimum essential medium (MEM; Thermo Fisher Scientific Inc., Waltham, MA, USA) containing 10% fetal bovine serum (FBS; Thermo Fisher Scientific Inc.) and 100 U/mL penicillin/streptomycin (10% α-MEM) on a 10 cm culture dish. The non-adherent cells were cultured with 30 ng/mL M-CSF in 10% α-MEM on a 10 cm Petri dish. After 3 days, the adhered cells were harvested and used as bone marrow-derived macrophages (BMMs). The BMMs were cultured with 10 ng/mL RANKL (R&D Systems, Minneapolis, MN, USA) and 30 ng/mL M-CSF in 10% α-MEM for 4 days in the presence of the vehicle (0.1% dimethyl sulfoxide [DMSO]) or EFV.
3. TRAP staining
The adherent cells were fixed with 10% formaldehyde for 5 min, permeabilized with 0.1% Triton X-100 for 10 min, and incubated with a tartrate-resistant acid phosphatase (TRAP)-staining solution (Sigma-Aldrich, St. Louis, MO, USA) at room temperature for 10 min. The TRAP-positive cells stained red and stained cells with 3 or more nuclei were counted as mature osteoclasts.
4. Cytotoxicity assay for extracts of Flavoparmelia sp.
BMMs were cultured with 30 ng/mL M-CSF in 10% α-MEM in the presence of the vehicle (0.1% DMSO) or EFV. After 3 days, the cell viability was assessed using a cell counting kit-8 (CCK-8; Dojindo Molecular Technologies, Kumamoto, Japan) according to the manufacturer's protocols.
5. Real-time polymerase chain reaction (PCR)
Real-time PCR was performed, as described elsewhere.[15] BMMs were cultured with 10 ng/mL RANKL and 30 ng/mL M-CSF in 10% α-MEM for the indicated days in the presence of vehicle (0.1% DMSO) or EFV. The primer sets for real-time PCR were designed (Table 1) using the online primer3 program.[16] The total RNA was obtained using the TRIzol reagent (Thermo Fisher Scientific Inc.) according to the manufacturer's protocol. First-strand cDNA was modified using a moloney murine leukemia virus cDNA Synthesis kit (Enzynomics, Daejeon, Korea) according to the manufacturer's instructions. Quantitative PCR (qPCR) was performed using the TOPreal qPCR 2×PreMIX (Bio-Rad, Hercules, CA, USA) in a Real-Time PCR Detection System (Bio-Rad). The relative levels of the tested genes were normalized to the level of glyceraldehyde-3-phosphate dehydrogenase and the data were analyzed using the 2−ΔΔCT method.[17]
6. Western blot
Western blotting was performed, as described previously.[18] BMMs were incubated in the same manner as real-time PCR assays. The cells were washed with phosphate-buffered saline and lysed in a lysis buffer (50 mM Tris-HCl, 150 mM NaCl, 5 mM ethylenediaminetetraacetic acid, 1% Triton X-100, 1 mM sodium fluoride, 1 mM sodium vanadate, and 1% deoxycholate) supplemented with 1 mM phenylmethylsulfonyl fluoride (Bio Basic Inc., Amherst, NY, USA). The lysates were centrifuged at 20,000×g for 13 min at 4℃ and the supernatant containing the proteins was collected. The proteins were subjected to 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to a polyvinylidene difluoride membrane (Millipore Corporation, Billerica, MA, USA). The membranes were blocked with 5% skim milk for 1 hr at room temperature and incubated overnight at 4℃ with the primary antibody. They were then incubated with the secondary antibody conjugated to horseradish peroxidase for 2 hr at room temperature. The membranes were developed with CLAROTM Mucho (Bio-D, Gwangmyeong, Korea) using a LAS-4000 luminescent image analyzer (Fuji Photo Film Co. Ltd., Tokyo, Japan).
7. Bone pit formation assay
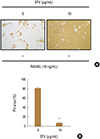
A bone pit formation assay was performed, as described elsewhere.[19] BMMs were seeded on an Osteo Assay plate (24 well plate; Corning, Tewksbury, MA, USA) at a density of 3×105 cells/well and cultured with 10 ng/mL RANKL and 30 ng/mL M-CSF in the presence of vehicle (0.1% DMSO) or EFV. After 4 days, the cells were removed completely with 5% sodium hypochlorite for 5 min, and the pit area was then observed by optical microscopy (magnification, ×50; Leica Microsystems, Wetzlar, Germany) and measured by ImageJ software (National Institutes of Health, Bethesda, MD, USA).
RESULTS
1. The methanol extract of Flavoparmelia sp.
HPLC analysis of the methanol extract of Flavoparmelia sp. showed that galbinic acid was the main component of the extract (Fig. 1). The retention time (Rt=2.242) and the maximum absorption wavelengths (λmax) of the UV-spectra (the insert in Fig. 1) of the peak confirmed that the compound was well matched with galbinic acid.
2. EFV inhibits RANKL-mediated osteoclastogenesis
The potential role of EFV in osteoclastogenesis was evaluated by examining the effects of EFV on the ability of RANKL to differentiate BMMs. BMMs were cultured with 0.1% DMSO (vehicle) or EFV (0, 1, 3, and 10 µg/mL) for 4 days in the presence of 10 ng/mL RANKL and 30 ng/mL M-CSF. Red stained TRAP-positive cells were induced by RANKL, but EFV reduced this induction (Fig. 2A). Moreover, EFV inhibited the number of TRAP-positive MNCs (nuclei ≥ 3) in a dose-dependent manner and the formation of MNCs was inhibited almost completely at 10 µg/mL (Fig. 2B).
3. EFV had no cytotoxic effect on BMMs
To determine if the anti-osteoclastogenesis efficacy of EFV is due to cytotoxicity, cell viability analysis was performed with a CCK-8 in BMMs. The BMMs were incubated with 0.1% DMSO (vehicle) or EFV (0, 1, 3, and 10 µg/mL) for 3 days in the presence of 30 ng/mL M-CSF. EFV had no cytotoxic effects on the BMMs at the indicated concentration in this study (Fig. 2C).
4. Effects of EFV on RANKL-induced gene expression
The effects of EFV on gene expression of NFATc1, a major transcriptional factor of osteoclast differentiation, were examined. Real-time PCR showed that RANKL gradually increased the mRNA level of NFATc1, but EFV decreased the transcriptional level of NFATc1 significantly (Fig. 3A). Moreover, EFV also reduced the transcriptional levels of TRAP, dendritic cell-specific transmembrane protein (DC-STAMP), and cathepsin K, the osteoclast differentiation marker genes regulated by NFATc1 (Fig. 3B–D).
5. EFV inhibited RANKL-induced protein expression of NFATc1
In the previous experiment, EFV reduced the mRNA expression of NFATc1. Hence, this study examined whether EFV affected the protein level of NFATc1 by western blotting. The protein level of NFATc1 was increased significantly by RANKL but was reduced dramatically by EFV (Fig. 4). This suggests that EFV inhibits the translational expression of NFATc1 and suppresses osteoclast formation.
6. Effects of EFV on RANKL-mediated bone resorptive activity of osteoclasts
Experiments were conducted to determine if the osteoclast formation inhibited by EFV also affects bone resorption. The osteoclasts formed wide pit areas on the bone slice, but EFV reduced the pit areas markedly (Fig. 5).
DISCUSSION
Lichens are a complex organism, in which algae or cyanobacteria (photobionts) live among the filaments of various fungi (mycobiont).[11] In addition, a lichen thallus often contains diverse assemblages of microfungi and microorganisms.[20] The combination of these various organisms allows the lichen to generate and use various secondary metabolites,[12] such as alkaloids,[21] peptides,[22] and terpenes.[2324] Their metabolites have been studied to discover new bioactive compounds [2526] and their biological activity is also being revealed.[272829]
Osteoporosis is a representative geriatric disease that is becoming a major concern because of the aging population. The disease results in an increased risk of bone fractures by lowering the strength of the bone because the number and activity of osteoclasts are increased due to a range of causes.[30] In addition, investigations have shown that the number of fracture patients due to osteoporosis and the cost of treatment are increasing.[313233] Therefore, it is necessary to study various means of preventing and treating bone diseases, such as osteoporosis. Osteoclasts are MNCs arising from hematopoietic stem cells/macrophage lineage. Their differentiation and function is controlled by M-CSF and RANKL, which are produced in mature osteoblasts and stromal cells.[734] M-CSF is involved in the growth, survival, proliferation, and differentiation of both hematopoietic and non-hematopoietic cells.[3536] In particular, it promotes the expression of the RANKL receptor, RANK, in osteoclastic progenitor cells.[37] RANKL is the most important cytokine during the osteoclast differentiation process.[1037] The RANKL-RANK signaling pathway activates NFATc1. As a result, the osteoclast precursor is differentiated into TRAP-positive MNCs (mature osteoclast).
In vitro screening experiments with the extract of various lichens extracts on RANKL-mediated osteoclast differentiation were performed prior to this study. The results showed that EFV inhibited the osteoclast differentiation. Subsequently, this study examined the inhibitive mechanism of EFV against osteoclast formation. First, EFV was treated at concentrations of 0, 1, 3, and 10 µg/mL during osteoclast differentiation to select the optimal concentration of EFV in this study. EFV significantly reduced osteoclast formation at concentrations ≥3 µg/mL and did not exhibit cytotoxicity even at concentrations of 10 µg/mL. The cells were treated with EFV at a concentration of 10 µg/mL, which showed optimal efficacy in the experiment, and then examined by real-time PCR and western blotting to identify the anti-osteoclastogenic mechanism.
NFATc1 is a master transcription factor on osteoclastogenesis and is activated by the RANKL signaling pathway. In this study, EFV inhibited RANKL-mediated mRNA and the protein expression level of NFATc1. In addition, EFV reduced the levels of TRAP, DC-STAMP, and cathepsin K expression, which are osteoclast formation and activation-related molecules, by decreasing NFATc1.[3738394041] And we confirmed that RANKL-induced bone resorption was decreased by EFV in vitro. This means that EFV inhibits and reduces the formation and activity of osteoclasts. Overall, these results suggest that EFV had a substance that affected osteoclast differentiation and function, and this substance reduced the expression of NFATc1, a key molecule during osteoclastogenesis.




 XML Download
XML Download