

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
More than 3 billion people suffer from allergies (i.e., asthma, atopic dermatitis and rhinitis) worldwide. Several protein allergens in the environment could induce allergic immune responses through activation of their functional components, including proteases.123
Protease-activated receptors (PARs) are activated by proteases. Although PAR2 reportedly mediates the T helper 2 (Th2)-immune response in allergic diseases, the exact underlying mechanism is poorly understood.45 In addition, thymic stromal lymphopoietin (TSLP) is produced by epithelial cells and triggers Th2-type inflammation by proteases.5 Moreover, PAR2 induces TSLP activation in response to serine proteases.5 The body is protected against the inflammation caused by proteases on epithelial cells due to the mucosal barrier (e.g., lung and skin). However, it is unclear whether these mechanisms are involved in allergic human diseases.
Reactive oxygen species (ROS) have recently been reported as an important factor influencing the development of allergic diseases.678 ROS also induce the Th2 response through the oxidization of lipids on epithelial cells, which in turn is mediated by the toll-like receptor 4 (TLR4) and the adaptor protein TIR-domain-containing adapter-inducing interferon (IFN)-β.9 Although the relationship between ROS and PAR2 has been addressed in a study on allergic asthma,10 the exact mechanism remains unclear.
In the present study, we aimed to assess the mechanism underlying the induction of allergic asthma by a cockroach allergen extract that includes protease. First, we evaluated the PAR2-ROS-TSLP-related initiation of inflammation and mucosal tight junction degradation caused by protease in the mouse airway in vivo and in human bronchial epithelial cells (A549 cells) in vitro. Secondly, the PAR2-antagonist (PAR2-ant) and anti-oxidants were used to block the initiation of inflammation in a mouse model of asthma and in A549 cells. Our findings could suggest whether alternative new treatments with PAR2-ant or anti-oxidants could be considered for allergic diseases.
MATERIALS AND METHODS
German cockroach extract (GCE) preparation
GCE was prepared as previously described.9 Fifty grams of frozen German cockroaches were homogenized in liquid nitrogen and defatted in 200 mL of ethyl ether and 200 mL of ethyl acetate. Extractions were performed with slow stirring at 4°C overnight in phosphate buffer saline (PBS) containing 6 mmol/L 2-mercaptoethanol and 1 mg/mL 1-phenyl-3-(2-thiazolyl)-2-thiourea to prevent melanization. The extract was then centrifuged at 10,000 ×g for 30 minutes at 4°C, and the supernatant was finally filtered through 0.2-μm filters. The endotoxin from the supernatant was removed by the Detoxi-Gel™ Endotoxin Removing Gel (Pierce Biotechnology, Rockford, IL, USA). The endotoxin from the GCE was removed and measured to be below 0.01 EU/mL by the chromogenic Limulus Amebocyte Lysate test (Lonza, Walkersville, MD, USA). The GCE contained 0.11 U/mg Bla g1.
ROS levels
The ROS levels were measured in human bronchial epithelial cells (A549) (Costar, Cambridge, MA, USA) in vitro and in the mouse lung in vivo. By using the OxiSelect ROS Assay Kit (Cell Biolabs, San Diego, CA, USA), ROS levels on A549 cells were measured according to the manufacturer's guidelines. In brief, the attached cells seeded on a black 96-well plate (1 × 105 cells/well) were pretreated with washing buffer and then treated with 100 μL of the cell permeable fluorogenic probe 2′, 7′ dichlorodihydrofluorescein diacetate (DCFH-DA, Sigma-Aldrich, St Louis, MO, USA) in Dulbecco's modified eagle medium (DMEM, Sigma-Aldrich) media for 1 hour at 37°C. After the cells were washed 3 times with Dulbecco's PBS, GCE (100 μg/mL) with or without PAR2-ant (NPS-1577, 200 μM; NeoPharm Pharmacy, Daejeon, Korea) was added to the DMEM media. Pretreatment with PAR2-ant was ensured for 30 minutes prior to GCE treatment. To quench ROS generation, 2 mM of N-acetyl-L-cysteine (NAC; Sigma A9165; Sigma-Aldrich, St Louis, MO, USA) was added. The plates were read at excitation and emission wavelengths of 450 and 520 nm, respectively, every 30 minutes for 2 hours by using a VICTOR™ X3 Multilabel Plate Reader (PerkinElmer, Santa Clara, CA, USA).
By using immunohistochemistry (IHC), the expression of ROS-related enzyme glutathione S-transferase 1 (GSTP1) was measured in the mouse lung in vivo (described in IHC section).
Cell viability assay
Cell viability was determined by using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. In brief, A549 cells at the logarithmic growth phase were randomly seeded onto 96-well culture plates at a density of 1 × 103 cells/mL, and were cultured with 100 μL DMEM media (supplemented with 10% fetal bovine serum [FBS]) in each well. After cell adhesion was achieved, the cells were incubated with various concentrations of GCE (0, 50, 100, 200 and 400 μg/mL) for 1, 2, 4 and 8 hours with or without PAR2-ant (200 μM) and NAC (2 mM). For the cell viability assay, 10 μL of MTT solution (Sigma-Aldrich; 5 mg/mL) was added to each well, and incubation was continued at 37°C for 4 hours. Following centrifugation at 3,000 rpm for 10 minutes, the supernatant was removed to obtain the formazan pellet. The pellet was then dissolved completely with 100 μL of dimethyl sulfoxide (DMSO). An enzyme linked immunosorbent assay (ELISA) plate reader was used to measure the absorbance at a wavelength of 570 nm, in order to determine the amount of pellet.
Real-time reverse transcription polymerase chain reaction (PCR) in the A549 cells
A549 cells were cultured in DMEM (Sigma-Aldrich) media and supplemented with 10% FBS (Sigma-Aldrich) and 1% antibiotics (penicillin/streptomycin; Invitrogen, Carlsbad, CA, USA) under standard tissue culture conditions. A total of 6 × 104 cells were seeded per well on 12-well plates, which were exposed to 100 μg/mL or 150 μg/mL (claudin-1) of GCE with or without 200 μM PAR2-ant and 2mM NAC for 2 hours.
Pretreatment with PAR2-ant was ensured for 30 minutes prior to GCE treatment. To confirm the PAR2 agonist (PAR2-ago) function of GCE, a positive control for the PAR2-ago (100 μM, SLIGKV-NH2; R&D Systems, Minneapolis, MN, USA) was also examined. To determine the optimal duration for each mediator (TSLP/claudin-1) on exposure to GCE, the expressions were measured at different time points (0, 2, 6, 8, 24, 36 and 48 hours) by using real-time PCR. At the optimal duration for each mediator, the other material (PAR2-ant, NAC and PAR2-ago) was added to the GCE-treated cells. Real-time PCR was performed with specific primers and probes using TaqMan Gene Expression Assays and TaqMan Gene Expression Master Mix (Applied Biosystems, Foster City, CA, USA) on an ABI 7500 system (Applied Biosystems, Piscataway, NJ, USA). The signals were normalized using glyceraldehyde 3-phosphate dehydrogenase (GAPDH).
Mouse model and treatment
Female BALB/c mice weighing 20–25 g (4 weeks of age) were purchased from OrientBio (Seongnam, Korea) and cared for and used in accordance with the guidelines of the Institutional Animal Care and Use Committee at Asan Medical Center and Ulsan University College of Medicine. The mice were intranasally sensitized to GCE (100 µg in PBS) on days 1, 3, 5, 8, 10 and 12, whereas PAR2-ant (0.02% in 1% DMSO, intranasally) or NAC (100 mg/kg in PBS, intraperitoneally) was administered 30 minutes before GCE treatment on days 1, 3 and 5. Five mice were included in each group.
Clinical evaluations and analysis in vivo
The bronchial hyperresponsiveness (BHR) in response to inhaled methacholine (MeCh; Sigma-Aldrich) was measured in conscious, unrestrained mice by using a barometric whole-body plethysmograph (Buxco; EMKA Technologies, Paris, France). For bronchoalveolar lavage (BAL) fluid analysis, the airways were lavaged 2 times through a tracheal cannula with 1-mL aliquots of pyrogen-free saline warmed to 37°C. The recovered lavage fluid was pooled, and the cells were collected by centrifugation (5,000 rpm, 4°C and 5 minutes) and resuspended in 100 mL of cold PBS. The cells were stained with trypan blue to determine viability, and the total number of nucleated cells were counted using a hemocytometer. For differential BAL cell counts, cytospin preparations were made and stained with Diff-Quik (Sysmex, Takatsukadai, Japan). After the samples were coded, all cytospin preparations were evaluated by a single observer using an oil immersion microscope (magnification, × 1,000). At least 200 cells were counted per preparation, and the absolute number of each cell type was calculated.
Serum was obtained after BHR measurement, and 100 μL (1/10 dilution in carbonate-bicarbonate buffer) was added to each well of a 96-well plate. An immunoglobulin E (IgE)-specific ELISA was used to determine the total IgE levels in the serum by using matching antibody pairs (eBioscience, San Diego, CA, USA) according to the manufacturer's instructions.
For the histological evaluation of lung tissue, the left lung of each mouse was embedded in paraffin, sectioned to a thickness of 5 μm, and stained with hematoxylin and eosin to assess eosinophilic infiltration. Inflammation was scored by 2 independent investigators in a blind manner. The degree of peribronchial and perivascular inflammation was evaluated on a subjective scale of 0–3.
To measure the levels of inflammatory cytokines (interleukin [IL]-4, IL-13, and IFN-γ), whole-lung RNA was extracted using RNeasy (Qiagen, Valencia, CA, USA), and real-time PCR was performed with specific primers and probes using TaqMan Gene Expression Assays and TaqMan Gene Expression Master Mix (Applied Biosystems) on an ABI 7500 system (Applied Biosystems). Signals were normalized using GAPDH.
IHC
Sections from formalin-fixed, paraffin-embedded tissue blocks of lung were cut (5-μm thickness) and mounted on glass slides. IHC was conducted using an automated slide preparation system Benchmark XT (Ventana Medical systems Inc, Tucson, AZ, USA). Deparaffinization, epitope retrieval and immunostaining were performed according to the manufacturer's instructions by using cell conditioning solutions and the BMK ultraVIEW diaminobenzidine detection system (Ventana Medical Systems). Lung sections were stained with TSLP (1:700 dilution, ab115700; Abcam, Cambridge, United Kingdom), GSTP1 (1:800 dilution, GTX112695; GeneTex, Irvine, CA, USA), and claudin-1 (1:25 dilution, 71-7800; Invitrogen). Positive signals were amplified by using ultraVIEW copper, and sections were counterstained with hematoxylin and bluing reagent.
Determination of immunohistopathological staining intensity
Immunohistopathological staining was objectively analyzed by using a computer imaging program (ImageJ® 1.41; http://rsb.info.nih.gov/ij/).
RESULTS
GCE induces inflammation and degrades claudin-1 via a PAR2- and ROS-mediated mechanism in human bronchial epithelial cells (A549 cells)
DCFH-DA analysis indicated that GCE treatment generated ROS after 2 hours in A549 cells, whereas PAR2-ant and NAC treatment significantly suppressed ROS generation (Fig. 1A and B). This suggests that GCE induces ROS generation, which is mediated via PAR2.
 | Fig. 1GCE induces inflammation and degrades tight junctions in human bronchial epithelial cells (A549 cells). The 2′, 7′-DCF intensity is significantly increased following GCE exposure and suppressed following PAR2-ant and NAC treatment in A549 cells (1 hour) (A) and 2 hours (B). The expression of TSLP is significantly increased following GCE and PAR2-ago treatment and suppressed following PAR2-ant and NAC treatment in A549 cells after 2 hours (C). The expression of claudin-1 is significantly decreased by GCE and PAR2-ago treatment, but is significantly restored by PAR2-ant and NAC treatment in A549 cells (D). (A, B) Fluorescence intensity measurement at wavelengths of 450 nm and 520 nm; (C, D) real-time polymerase chain reaction (Taqman method).GCE, German cockroach extract; NAC, N-acetylcysteine; DCF, dichlorofluorescein; PAR2-ant, protease-activated receptor 2-antagonist; PAR2-ago, protease-activated receptor 2 agonist; TSLP, thymic stromal lymphopoietin.
*P < 0.01 and †P < 0.001. The experiments were repeated 6 times.
|
To evaluate the ROS- and PAR2-related inflammatory mechanism in the A549 cells following exposure to GCE, we measured the levels of TSLP and claudin-1 using real-time PCR. The expression of TSLP was significantly increased by GCE or PAR2-ago treatment after 2 hours (Fig. 1C). However, the expression of claudin-1 was significantly suppressed by GCE or PAR2-ago treatment after 6 hours (Fig. 1D). Thus, it is likely that the expression of inflammatory cytokine TSLP and tight junction protein claudin-1 is mediated by PAR2 following exposure to GCE in A549 cells.
The expression of TSLP was significantly suppressed by PAR2-ant and NAC treatment (Fig. 1C), whereas the expression of claudin-1 was significantly increased following PAR2-ant and NAC treatment (Fig. 1D). Hence, the induction of inflammatory mechanisms involving the TSLP and tight junction protein degradation by GCE may be mediated by PAR2 and ROS.
GCE induces allergic inflammation, which is mediated by a PAR2 and ROS mechanism, in a mouse model of asthma
To confirm whether the PAR2 and ROS mechanism was involved in the response to GCE in vivo in a similar manner to that noted in vitro, a mouse model of asthma was established using GCE. The phenotypes of allergic asthma including BHR, total cell count in BAL fluid, total IgE levels, histopathology and lung inflammatory score were significantly increased following GCE treatment (Fig. 2). Differential cell counts indicated eosinophilic and neutrophilic inflammation in the BAL fluid (Fig. 2C). The phenotypes were significantly suppressed by PAR2-ant and NAC treatment. This suggests that the allergic inflammation in the mouse model was mediated via the PAR2 and ROS mechanism.
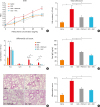 | Fig. 2GCE induces allergic inflammation in the mouse model of asthma. (A) BHR is significantly increased following GCE treatment and suppressed following PAR2-ant and NAC treatment. ‡P < 0.001 compared to saline. (B) The total number of cells in bronchoalveolar lavage fluid is also significantly increased following GCE treatment and suppressed following PAR2-ant and NAC treatment. (C) The numbers of macrophages, eosinophils and neutrophils are significantly increased following GCE treatment and suppressed following PAR2-ant and NAC treatment. (D) The total immunoglobulin E level is also significantly increased following GCE treatment and suppressed following PAR2-ant and NAC treatment. (E) Lung inflammation (hematoxylin and eosin staining 200 × magnification) and (F) peribronchial inflammation is significantly increased following GCE treatment and suppressed following PAR2 and NAC treatment.BHR, bronchial hypersensitivity; GCE, German cockroach extract; CE, cockroach extract; NAC, N-acetylcysteine; PAR2-ant, protease-activated receptor 2-antagonist.
*P < 0.05, †P < 0.01, and ‡P < 0.001. The experiments were repeated 5 times.
|
GCE induces Th2 inflammation in the lung, which is mediated by the PAR2 and ROS mechanism, in a mouse model of asthma
The level of Th2 cytokines IL-4 and IL-13 were significantly increased in the lung following GCE treatment, and were suppressed following PAR2-ant and NAC treatment (Fig. 3A and C). In contrast, the expression of Th1 cytokine IFN-γ was significantly suppressed by GCE treatment and increased following PAR2-ant and NAC treatment (Fig. 3B). This suggests that GCE induces Th2 inflammation in the lung, which is mediated by the PAR2 and ROS mechanism, in a mouse model of asthma.
 | Fig. 3GCE induces T helper 2-allergic inflammation in a mouse model of asthma. The levels of IL-4 and IL-13 are significantly increased following GCE treatment and suppressed following PAR2-ant and NAC treatment in the lung (A and C). Moreover, the level of IFN-ν is significantly decreased following GCE treatment and increased following PAR2-ant and NAC treatment (B). Data were measured using real-time polymerase chain reaction.GCE, German cockroach extract; IL, interleukin; IFN, interferon; PAR2-ant, protease-activated receptor 2-antagonist; NAC, N-acetylcysteine.
*P < 0.05; †P < 0.01. The experiments were repeated 5 times.
|
GCE induces ROS-TSLP-related inflammation in the lung, which is mediated by PAR2, in a mouse model of asthma
The expression of ROS-related enzyme GSTP1 in the lung was significantly decreased following GCE treatment. IHC of the lung indicated that the decreased GSTP1 levels had recovered following PAR2-ant and NAC treatment (Fig. 4). Moreover, the expression of TSLP in the lung appeared to be regulated via the PAR2-ant and NAC mechanism, as per the findings of IHC examination (Fig. 5). Thus, the ROS generation by GCE treatment was followed by an increase in TSLP expression, which was mediated by PAR2, in the lung of a mouse model of asthma.
 | Fig. 4GCE significantly suppresses GSTP1 expression in a mouse model. GSTP1 expression is significantly increased by PAR2-ant and NAC treatment in the lung, as indicated by immunohistopathology (immunohistochemistry, 200 × magnification) (A) and intensity quantification (B).GCE, German cockroach extract; GSTP1, glutathione S-transferase 1; NAC, N-acetylcysteine; PAR2-ant, protease-activated receptor 2-antagonist.
*P < 0.01; †P < 0.001. The experiments were repeated 5 times.
|
 | Fig. 5GCE significantly increases TSLP expression in a mouse model. TSLP expression is significantly suppressed by PAR2-ant and NAC treatment in the lung, as indicated by immunohistochemistry (200 × magnification) (A) and intensity quantification (B).GCE, German cockroach extract; TSLP, thymic stromal lymphopoietin; NAC, N-acetylcysteine; PAR2-ant, protease-activated receptor 2-antagonist.
*P < 0.001. The experiments were repeated 5 times.
|
GCE causes disruption of the tight junction protein in the lung, which is mediated by a PAR2- and ROS-related mechanism, in a mouse model of asthma
Claudin-1 expression was disrupted by GCE treatment in the lung, but recovered following PAR2-ant and NAC treatment, as noted on IHC examination (Fig. 6). This suggests that GCE induces tight junction degradation, which is mediated by a PAR2- and ROS-related inflammatory mechanism.
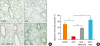 | Fig. 6GCE significantly disturbs claudin-1 expression in a mouse model. Claudin-1 expression is significantly restored by PAR2-ant and NAC treatment in the lung, as indicated by immunohistochemistry (200 × magnification) (A) and intensity quantification (B).GCE, German cockroach extract; PAR2-ant, protease-activated receptor 2-antagonist; NAC, N-acetylcysteine.
*P < 0.01; †P < 0.001. The experiments were repeated 5 times.
|
DISCUSSION
In the present study, we found that ROS-TSLP-related inflammation was induced by GCE exposure via PAR2 in human bronchial epithelial cells and a mouse model of asthma. Moreover, we observed that tight junction proteins could be affected in allergic inflammation via PAR2 in a mouse model of asthma. Although asthma is regarded as an airway inflammatory disease that is driven by a skewed Th2 mechanism, the process of initiation of the disease on the surface of respiratory epithelial cells remains unclear. Protease allergens reportedly elicit airway reactions by triggering innate immunity receptors, such as PARs, and thus activating epithelial cells, mast cells and dendritic cells (DCs), which in turn induce the additional release of mediators.11 In patients with bronchial asthma, PAR2 expression is increased on the surface of respiratory epithelial cells.12 Hence, blocking of PAR2 could be considered as an option for the treatment of asthma. Although some studies have indicated the presence of a PAR2-related mechanism in the mouse model,1013 no studies have shown any specific association among the mediators that could play an important role in the development of asthma.
The present study showed that the protease-containing allergen GCE first activated PAR2, and then induced ROS generation in A549 cells and in the mouse model. The present study also showed PAR2 could be one of the possible mechanism of allergic inflammation. ROS play a central role in orchestrating Th2 inflammation, by inducing the formation of oxidized lipids that trigger the TLR4 and toll or IL-1 receptor domain-containing adaptor-inducing IFN-β-mediated induction of TSLP in epithelial cells.9 In the present study, the expression of TSLP was regulated by PAR2-ant or anti-oxidants, and ROS generation was mediated by PAR2. Hence, TSLP appears to be induced by the ROS generated by PAR2 activation, consistent with the findings of a previous study.9
Mutation of the barrier protein filaggrin is a critical factor in the pathogenesis of atopic dermatitis.1415 Although filaggrin is not expressed in the lower airway respiratory epithelium,16 the barrier function of the airway epithelium is impaired in asthma, thus indicating a shared common underlying pathogenic mechanism.17 In the present study, the expression of claudin-1 was significantly decreased following exposure to GCE or PAR2-ago in A549 cells, and was also decreased in the lung of a mouse model sensitized to GCE. Thus, the findings confirm that the tight junction protein claudin-1 is closely related to the development of asthma. The expression was also regulated by PAR2 and ROS in vitro and in vivo, suggesting that tight junction proteins could be degraded through the activation of PAR by proteases in allergens and via the generation of ROS.
The initiation of sensitization to allergens in the airway is preceded by their uptake and processing by a subpopulation of mucosal DCs, followed by the presentation of specific peptide epitopes to naïve T cells in association with major histocompatibility class II.18 Protease enzymatic activity of allergens that could promote intraepithelial DCs functions for allergic reactions by cleaving the tight-junction proteins.19 The activation of DCs could induce Th2 cell responses via epithelial cell-derived factors such as TSLP, which conditions DCs to induce Th2 cell responses.2021 Thus, allergen source-derived proteases compromise epithelial barrier function by degrading tight junction proteins, thus facilitating allergen delivery across the epithelial layers.17 The above mentioned theory is summarized in Supplementary Fig. S1.
In conclusion, GCE activates PAR2 and generates ROS, which degrade tight junction proteins and facilitate the penetration of allergens to induce TSLP-Th2 inflammation in airway epithelial cells in vitro and in the lung of a mouse model. Thus, the blocking of PAR2 is an alternative treatment option for allergic inflammation.
 XML Download
XML Download