

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
The effects of environmental changes on our planet (for example climate change) have become more evident in governments and public health authorities during the last few years. Complex, multifactorial diseases such as allergic disease have been increasing during the last decade and there are reasons to believe that global and regional changes could be important determinants.1 Since symptoms of allergic disease are induced and triggered by allergens, it is possible that climate and other environmental influences on asthma prevalence are due to modifications of the persistence, quality and intensity of allergen exposure,2 leading to an increase in allergic responses and clinical symptoms. House dust mite (HDM) allergens are important risk factors for asthma34; indeed, they are the main risk factor for this disease in tropical regions.56 However, in contrast to pollen and fungal allergy,789 the impact of global environmental changes on natural HDM growth and allergen production has not been completely evaluated. Also, it remains to be defined if there is an increase in sensitization to HDM allergens and its possible relationship with global or local environmental changes. To better understand these problems, this review starts with a brief presentation of immune responses to allergens and then explores: (1) the epidemiological trends toward HDM sensitization and mite-induced allergic symptoms; (2) the current and projected influence of environmental changes on HDM exposure; (3) the role of adjuvants of the immunoglobulin E (IgE) response to HDMs; and (4) the potential role of the genome and epigenome in adaptation to environmental changes. Regarding the potential and demonstrated impact of climate change on allergic diseases associated to pollen allergen exposure, several reviews are available.110
GENERAL ASPECTS OF IMMUNE RESPONSES TO HDM ALLERGENS
Adaptive immunity pathways
In genetically susceptible individuals, immune responses to allergens are mainly based on specific IgE and type 2 cytokines, such as interleukin (IL)-4, IL-5 and IL-13. Specific IgE primary responses to allergens are exclusively produced through the adaptive pathways of immunity. After sensitization, re-exposure to allergens initiates the IgE-dependent inflammatory cascade, which involves additional cells and ILs that actively participate in inflammatory reactions. Under shared environments and levels of exposure, only a small proportion of people have this response; therefore, genetic factors have been extensively investigated to conclude that they play an important role in Th2 immunity. It is well known that environmental conditions determine not only the adaptive Th2 response to allergens, but also the inception of allergic diseases.1112 Although the association of HDM allergen concentration with IgE sensitization is not always positive,1314 theoretically high environmental levels of these allergens will increase the probabilities that genetically susceptible individuals inhale them and become sensitized. The following factors, sometimes acting concomitantly, modulate the IgE primary response and sensitization process: the level of exposure (which in turn depends on protein production by the mite, permanence in the house dust and the air, and effects of other enzymes upon the allergen), persistence of exposure, age of exposed individuals, boosting the IgE responses by conditions such as air pollution or helminth infections, and stimulation of Th1 response by bacterial and other products. The impact of environmental changes on these factors might modify the frequency and strength of allergen sensitization; therefore, some of them will be assessed in this review.
Innate immunity pathways
Historically, the word allergen has been associated with a specific-IgE inducer or reactor. Thus, non-infectious inducers and IgE-binding components are generally considered allergens, which does not mean they have the same pro-inflammatory properties. Allergenic activity can be increased if the allergen can induce inflammation by other pathways, in addition to specific IgE. In fact, several innate non-IgE mediated inflammatory mechanisms have been reported, including the ability to bind adjuvants or to stimulate the innate immunity via toll-like receptors and other receptors of the bronchial epithelium.1516171819 Der p 1 and Der p 2 are good examples of molecules with high allergenic activity probably because of their ability to stimulate both the innate and adaptive pathways of allergic responses.
EPIDEMIOLOGICAL TRENDS OF HDM SENSITIZATION
Since in the tropics HDM allergy is so frequent, it is worth investigating if recent environmental changes are affecting the sensitization there. The tropics are regions of the Earth surrounding the Equator, delimited in latitude by the Tropic of Cancer in the Northern Hemisphere and the Tropic of Capricorn in the Southern Hemisphere. In some regions, the tropical environment can extend beyond these parallels by the effect of oceanic streams and other geographical characteristics. These conditions provide the ideal warm and humid environment (25°C to 30°C and a relative humidity of 70%) for the growth and multiplication of HDMs. There are differences in the distribution of HDM species; for instance, Blomia tropicalis is prevalent in the tropics and subtropics,20212223242526 while Dermatophagoides farinae and Dermatophagoides pteronyssinus reproduce at cooler temperatures and are inhabitants of homes worldwide.27 Lower winter temperatures in combination with heated homes reduce D. pteronyssinus rather than D. farinae because the latter is more resistant to reduced levels of relative humidity (RH) and can resist periods of drought, thereby it is more frequently observed in homes from the northeastern regions of North America, Northern Europe and Korea.28
Allergic diseases have increased worldwide, especially in tropical regions of Asia-Pacific, Africa and Latin America. Several hypotheses have been proposed to explain these trends,29 but one interesting observation is increased IgE sensitization to HDMs in communities where allergies have been traditionally low prevalent. IgE reactivity to HDM allergens has been evaluated since the early 1990s in different regions of the world. Differences in study design, definition of sensitization (skin prick test vs.in vitro IgE determination),30 age range of patients (pediatric vs. adults), geographic location (urban vs. rural) and the clinical definitions of patients, precluded a systematic comparison on IgE sensitization frequencies over time. IgE sensitization to B. tropicalis among patients with respiratory allergies and asthma was already high (> 70%) in Brazil, Colombia, Cuba and Singapore in the early 1990s as it is so nowadays. However, the most remarkable changes in the characteristics of HDM fauna and sensitization rates have been detected in the subtropics and temperate areas including Taipei City in Taiwan,31 Southern China and India.32 It is possible that economic development influences these trends as suggested by the differences in HDM sensitization between rural and urban areas.
Due to their rapid industrialization, it is in urban developed regions of Asia where the increase in HDM sensitization has been more pronounced. Figures of HDM sensitization in China, Taipei, Indonesia and Korea reach about 80% to 90% in patients with respiratory allergies.33 IgE sensitization to HDMs goes in parallel with the level of urbanization as shown in a study reporting positive IgE to D. pteronyssinus in 49.7% of individuals in rural Taiwan compared to 60% to 80% in Taipei City. Similarly, Guangzhou (China) underwent a process of urbanization in recent years, which together with a subtropical climate provided appropriate conditions for HDMs to grow and reproduce, led to 80% frequency of HDM sensitization and potentially influenced the increasing trends of allergic diseases in that country.34
Increased rates of HDM sensitization are also prominent in urban areas of Africa, acting as a major risk factor in children living in affluent localities.35 HDMs are also important sensitizing allergens in deserted places, with sensitization rates of 76% in asthmatic patients and 62.3% of rhinitis patients from Kuwait.3637 In contrast, the rates of HDM sensitization have decreased from 63.2% to 45.9% between 1992 and 2002 in Zagreb, while ragweed sensitization has significantly increased.38
In addition to increasing rates, in the last 2 decades there has been a shift in the patterns of sensitization toward HDM species. In Singapore, B. tropicalis was the most prevalent HDM in 1999,21 but a study in 2005 revealed that Dermatophagoides species induced higher rates of sensitization in Singaporean children.39 However, it has also reported a remarkable bias toward IgE sensitization to HDM allergens in Singaporean adults with more equilibrated rates between Blomia and Dermatophagoides.6 For unclear reasons, Tyrophagus putrescentiae40 and B. tropicalis are not so predominant as before in developed Asian cities; probably, the decrease in hay, mold and agricultural societies is shifting the conditions toward the predominance of Dermatophagoides species. Similarly, in rural central Taiwan, the sensitization is more frequent to B. tropicalis than to D. pteronyssinus, which is predominant in urban Taipei.41
Differences in sensitization frequencies and the type of HDMs is recognized by young and old age groups, suggesting that environmental changes in the last decades may be affecting mite biology and thereby sensitization rates. For example, elderly subjects in Taiwan (over 70 years of age) have higher sensitization rates to T. putrescentiae (greater than that to D. pteronyssinus) compared to younger adults (< 40 years old).42 Other studies showed that this mite was the third most common in Korean homes,43 although its relevance seems to have changed overtime4445 and largely depends on the geographical region.46 Moreover, a comparative analysis of the sensitization rates to HDMs from 1980 to 2010 in Korea found that younger age groups (10-20 years old) had HDM sensitization rates of 70% compared to 30% in the group above 60 years old.47 In addition, skin prick test reactivity to D. pteronyssinus (43.2%) significantly increased in the 1990s compared to the 1980s (33.4%), but did not differ between the 1990s and the 2010s.47 HDM sensitization has also increased since 1994 in secondary schoolchildren from Guangzhou, China,4849 although another study in the same region reported no difference in the overall prevalence of HDM sensitization (up to 85%) at 10-year intervals from January 2005 to December 2014. This study also showed that HDM sensitization was higher in younger age groups (10-19 years, 91.4%) than those above 50 years (68.5%).50 The fact that younger age groups had more IgE reactivity toward HDM allergens cannot totally be explained by immunosenescence, suggesting new patterns of HDM sensitization in emerging economies during the previous years.
THE CURRENT AND PROJECTED INFLUENCE OF ENVIRONMENTAL CHANGES ON HDM EXPOSURE
Factors affecting HDM growth and survival
Temperature
Evidence supports an increase in global temperature (https://climate.nasa.gov/).51 The most immediate consequence is that tropical environments will expand to latitudes beyond the Tropics of Cancer and Capricornus; thereby, more people will get exposed to HDM allergens. Other regions in higher latitudes, albeit still temperate, will experience milder and more humid winters which might increase HDM exposure.52 The effects of global warming have been identified by studies showing that high temperatures are associated with increased asthma symptoms53 and with increased risk of repeated hospital admission in children with asthma.54 Moreover, average annual temperature was the main outdoor factor that correlated with higher mite concentrations.55 Since HDM species have different preferences for temperature,56 it could be anticipated that climate change can exert their selection in some places. Temperature is known to affect hatching of HDM eggs57 and HDM allergen production.58 It is possible that climate change might affect mite metabolism and that new allergens may become very relevant as sensitizing sources.
Increased temperature has also modified human lifestyles and made individuals spend more time indoors and use air conditioning which can in turn influence HDM exposure.
Humidity
Humidity is an important determinant of HDM growth and survival. According to the Intergovernmental Panel on Climate Change (IPCC, https://iaqscience.lbl.gov/cc-humidity), local increases and decreases in indoor air absolute humidity are expected as a result of climate change. Increased temperature can cause a rise in humidity, possibly influencing the growth and survival of HDMs. This effect could occur in temperate zones, where the increase in relative humidity during the whole year can augment the level and persistence of exposure. Humidity is a great limiting factor for HDM growth and has a profound influence on their reproduction.5960 Mites absorb vital moisture from the air through their exoskeletons and supra coaxial glands, requiring high air humidity to prevent excessive water loss.61 HDMs depend on water activity (Aw), which is the RH at a surface. Also, in the critical humidity, water intake by passive and active mechanisms compensates for water loss. The RH is defined as the amount of water vapor in a given volume of air at a given temperature, expressed as the percent of the maximum possible for that temperature. A decrease in ambient RH (which is paralleled by an Aw drop) affects HDMs not only in laboratory settings but also in their natural environments. Optimal RH values for HDM growth have been found between 70%–90%62; however, critical air humidity differs among mite species,63 for instance, 60%–65% for D. pteronyssinus, 47%–50% for D. farinae and 74%–80% for B. tropicalis.64 The environmental humidity affecting room microclimate is consequently the main reason why some species are predominant under different geographical conditions. For instance, D. pteronyssinus is more susceptible to desiccation than D. farinae. The lowest humidity/temperature HDMs can survive is 55%–74%/15°C–35°C; when humidity levels are less than 55%, dust mites will gradually dehydrate and die. This is the case for many homes where the winter is cold and dry, causing a low RH in the heated indoor air. Some studies in temperate areas suggest that maintaining an indoor RH of less than 51% during the humid summer season resulted in significant reductions in mite and allergen levels.65 Increased house humidity has been associated with increased prevalence of allergic and respiratory symptoms of asthma as well as IgE levels66 and is a well-recognized risk factor for HDM allergy worldwide.
Human activities have already increased the moisture content of the atmosphere; in accordance with the Clausius–Clapeyron relationship, rising global temperatures will increase humidity (HadISDH - gridded global land surface humidity dataset - version 4.0.0.2017f).51 By measuring “wet bulb” temperature, which reflects the combined effects of heat and humidity by draping a water-saturated cloth over the bulb of a conventional thermometer, new information has been obtained about this topic. A study on projected “wet-bulb” temperatures shows that in the Southeastern United States, wet bulb temperatures of 28.8°C are rare, but by the 2070s and the 2080s this condition could happen 25 to 40 days each year, being worse in Northern India, Central and Western Africa, Eastern China and South America. Also, by 2080 extreme wet bulb conditions could become 100 to 250 times more frequent in the tropics and increases of 10%–15% humidity are projected across Eastern US, Northeastern India, Eastern China and West Africa (https://www.clim-past.net/10/1983/2014/). A summary of the worldwide changes in surface relative humidity between 1973 and 2017 is presented in Fig. 1. Since HDMs have developed interesting humidity-oriented adaptations to survive in their natural environments (i.e. resistance to reduced RH or tolerance to desiccation), we can expect that anomalies in at least 5% in RH may impact HDM growth.
 | Fig. 1Changes in worldwide relative humidity from 1973 to 2017 as reported by the HadISDH - gridded global land surface humidity dataset - version 4.0.0.2017f. Maps were downloaded from https://www.metoffice.gov.uk/hadobs/hadisdh/.
|
If winters become milder, there will be more HDMs and possibly a shift in their species. For example, if the Northeast United States becomes warmer and more humid, such mites as Blomia tropicalis may extend their range northward in the United States. Changes in relation with humidity has already been detected in the study of Antens et al.,67 in which Der f 1 apparently became the most highly concentrated allergen in house dust between the beginning of the study in 1996 and 8 years later. HDM sensitization has also been observed in tropical areas of high altitude68 as well as in alpine temperate regions,69 so that it is feasible that with humidity changes some areas considered dry and at high altitude may become suitable for HDM distribution. Changes in humidity in northern latitudes will increase the numbers and species of HDMs. It is worth to mention that humidity has a critical influence on fungi and affect its interactions with HDM populations.7071 Fungi digest lipids from protein substrates, and provide mites with vitamins and sterols that facilitate the assimilation of food by HDMs.72 In addition, it has been found that HDMs ingest fungi.73 However, if RH increases over 90%, there is the disproportionately growth of fungi leading to the mortality of HDM nymphs and reducing the lifespan of adult mites.
The increasing rates of HDM sensitization in the Asia Pacific region and other developing areas of South America seem to be also associated with the rapid changes in urbanization and lifestyle.74 Residential environment improvements acquired with economic development provide a better habitat not only for humans but also for HDMs. Urbanization also introduced changes in behavioral aspects that lead humans to spend more time indoors, altogether promoting HDM exposure. A rise in humidity in indoor air can affect the growth of HDM and allergen production; thereby, factors modifying the control of the indoor environment such as air conditioning (AC), ventilation systems, and building materials will be critical in facilitating HDM exposure and sensitization. With increased population and energy costs, the human individual area in urbanized major cities tends to reduce. Building strategies have changed to make smaller and tighter homes, often resulting in higher indoor humidity and reduced indoor air quality. Cooking devices in poorly ventilated spaces create water vapor that also affects indoor air. Moreover, the use of AC is increasing, and some studies suggested that they remove water vapor and reduce indoor humidity. Indeed, maintaining indoor RH between 35% and 50% decreases HDM growth. Patients living in households without AC are at increased risk of mold sensitization and polysensitization75; however, it depends on the system and the characteristics of the household. Those with evaporative systems in low socioeconomic homes have shown to increase humidity and to promote dust mite growth.76 AC can also increase indoor dampness by leakage of water in walls and carpets as well as by creating condensation in walls and windows. The overall results of moisture and water accumulation may lead to HDM and mold growth.
Factors affecting HDM allergen production
As occurs in other living beings, mite metabolism and homeostasis largely depend on the environment; therefore, changes in diet, quality of air and water, temperature, and humidity or exposure to infectious agents could modify the level of gene expression of different cell components, some of them allergenic for humans. Several examples are given below (Table 1).
Table 1
Factors affecting HDM-allergen production

![]()
Proteases
HDMs have enzymes with protease activity, and those allergenic are most abundant.77 Group 1 allergens are cysteine protease (e.g. Der p 1), and groups 3, 6 and 9 are serine proteases with different substrate affinity. HDMs feed on sloughed skins as well as other sources such as commensal bacteria and fungi. Mite proteases seem to be mainly related to food digestion and are abundant in the digestive tract and fecal pellets.78 Mite speciation is markedly evident in protease gene content and expression.77 For example, cysteine proteases are 100 times more abundant in Dermatophagoides genus compared to other important mites including storage mites that feed on grains and dried fruits. Although B. tropicalis has relatively lower cysteine protease activity, serine protease activity is 10 times greater than observed in Dermatophagoides spp.79 Of note, even closely related mites, such as D. farinae and D. pteronyssinus, use different serine protease genes for the digestive process. D. pteronyssinus produces a collagenolytic enzyme, Der p 9, which is absent in D. farinae due to its low gene expression.7880 As general interpretation, protease expression in HDMs is fine-tuned with the nutritional composition of their natural feeding sources. In this sense, it is expected and also experimentally supported that mite diet may modify Der p 1 abundance.81
Among the projected adverse consequences of climate change is the increase of meteorological events such as hurricanes and cyclones. Regarding indoor air quality, a negative impact of these disasters is the growth of molds from flooding and water-damaged dwellings. As HDMs share their habitats with different fungal species and interact with them in different ways, fungus growth could increase HDM protease production, since they are required to degrade fungal cell walls.82 This could explain the fact that under experimental cultures, Der p 1 content in extracts prepared from mites grew in yeast-free medium were lower.81 The effect of temperature seems to act in the opposite way on controlled environments where only this factor is modified. During the exponential growth phase of HDM cultures, Der p 1 accumulated 1.38 times faster at 20°C than at 25°C.81
Lipid binding proteins
Lipid-binding proteins account for 20%–30% of HDMs and this biological property may be an adjuvant of allergenic activity.83 In addition, they activate Toll-like receptor (TLR)-2 or TLR-4 mediated pathways promoting inflammatory pathways in epithelial cells.15168485 Der p 2 has a strong affinity for lipopolysaccharide (LPS).84 Group 7 allergens are like mammalian LPS-binding proteins, although Der p 7 has affinity to another lipid ligand from bacteria and to the fungal lipopeptide polymyxin B.86 Due to their resemblance with innate immunity components from higher vertebrates, it is believed that they represent pathogen sensors in mites, as occurs in Drosophila melanogaster, where NPC2 proteins bind bacterial cell wall components and participate in immune signal pathways.87 Group 14 allergens include the vitellogenin/apoliphorin family. In bees, they are allergenic and display antimicrobial and anti-fungal activities related with their lipid binding function.88 Expression of vitellogenin is highly susceptible to external pressures; for example, in lipid-rich diets Der p 14 was found as the unique gene significantly overexpressed.89 Regarding the effect of climate change on lipid binding allergens, the best supported hypothesis is that a higher burden of fungi in indoor environments (due to higher air humidity) could increase the expression of certain lipid binding allergens considering that some fungal species are pathogenic for HDM. As observed in mammals, up-regulation of innate sensors after exposure to pathogens may also occur in mites. Batard et al.81 observed that Der p 2 expression was reduced on yeast-free medium. Temperature also increases group 2 expression; Der p 2 accumulates 1.41 times faster at 25°C than at 20°C.81
Chitin-related proteins
Chitin is a rigid polysaccharide polymer from the exoskeleton of fungi and arthropods, and it is also part of the peritrophic matrix that surround fecal pellets. It seems to modulate the immune response of mammals.90919293 Chitinases are hydrolytic enzymes required for the growth and morphogenesis of fungi and arthropods, including mites; however, in higher vertebrates, they protect against chitin-rich pathogens. Also, to digest fungi, mites require to degrade chitin. For the chitinase and chitinase-like allergen groups 15 and 18, their chitin-binding activity have been confirmed, but there is no experimental evidence about their chitinolytic functions.18 Assuming that an increase in humidity leads to higher burden of fungi in indoor environment,94 it may be possible that the expression of chitin-related proteins increases due to their roles in immunity and digestion.
Glutathione transferases
Three HDM allergens have been reported as mu-like glutathione transferases (GST). As observed in different invertebrates, exposure to toxic compounds increases GST gene expression. It has been observed that chronic exposure to diesel exhausted particles increases Der p 8 transcription as detected by reverse transcription polymerase chain reaction. In this context, it is possible that air pollution increases GST expression as observed with pollen allergens.95 This impact is also dependent on how indoor air quality is affected by external contamination. Another possibility is related to the increased risk of helminthiases reported in floodingareas,96 it has been reported that GST proteins may cross-react among mite and helminths.97
HDM structural proteins
Among invertebrates, several structural proteins have been found to be allergenic, showing a high degree of sequence homology. Best studied are the tropomyosins, a group of cross-reactive allergens from different sources: HDM, cockroaches, nematodes, mosquitoes and crustaceans.9899100 Different environmental changes may increase the risk of sensitization to tropomyosins. Aedes aegyptiis spreading to areas where it has previously been undetected. A parallel risk of flooding is the increase in pests such as cockroaches and helminths.96
FACTORS ADJUVATING THE IgE RESPONSE TO HDM
Air pollution
Air pollution has been one of the most detrimental environmental changes since industrial revolution; its characteristics and effects on increasing allergic sensitization and symptoms have been reviewed elsewhere.11 Several pollutants can induce bronchial inflammation and modify the immune responses,101102 (Table 2) sometimes increasing the Th2 allergic reactivity.103104 For example, co-exposure to DEPs can enhance the allergic response and HDM-induced airway hyperresponsiveness.105 In addition, residue oil fly ash, another component of the ultrafine fraction of particulate matter, also induces allergic pulmonary inflammation and acts as adjuvant of the allergic response to HDMs.106107 A significant increase in the transcription of allergens Der p 3, Der p 8 and Der p 21 was observed after exposing mites to a high concentration of diesel exhaustparticles.95 Moreover, the co-exposure to 0.1 ppm NO2 and Der p 1 significantly increased both IL-6 and IL-8 release,108 which suggests that air pollution can increase the inflammatory properties of some allergens. Indoor pollutants might also influence HDM allergenicity as reported that simultaneous exposure to HDMs and hexabromocyclododecane can enhance the antigen presentation and maturation/activation of dendritic cells.109
Table 2
Factors adjuvating the IgE response to HDMs

![]()
Helminth infections
Regarding allergic diseases, helminth infections have a dual effect: during severe infections they induce immunosuppression of the Th2 response and diminish allergic symptoms. In contrast, when they are light, with low parasitic load, they increase allergic responses.110 Helminth infections are going progressively controlled in most regions, even in developing countries, where they remain as public health problems. In many urbanized regions, improved hygienic measures have led to reduced parasite burden and this directly correlates with increased IgE sensitization to HDM allergens as detected by skin prick tests and in vitro IgE tests. The reduction of the immunomodulatory effects of parasitic infections (especially those by soil transmitted helminths) might boost IgE reactivity to HDMs. This means that the main global current immunologic effect of helminthiases is stimulating Th2 responses. The mechanisms include the high level of cross-reactivity between HDM and helminth (e.g. Ascaris lumbricoides) allergens9798 and non-specific boosting of HDM-specific IgE by still unknown helminth products.110111112 Deworming studies have shown that monthly anthelmintic treatment of children for 18 months caused an increase in the prevalence of skin test reactivity to HDMs in Venezuela113 and that treatment with praziquantel and mebendazole every 3 months for 30-months was associated with an increase in the incidence of skin reactivity to HDMs in Gabon.114 A more recent study also confirmed that positive skin test results to HDMs increased from 18.7% to 32.7% after albendazole treatment in Flores Indonesia.115 Interestingly, a 2-fold increase in allergen sensitization was observed in Ethiopian immigrants after 1 year in Israel, particularly for HDMs.116
THE ROLE OF THE GENOME AND EPIGENOME ON ADAPTATION TO ENVIRONMENTAL CHANGES
The environment is the great modifier of genome and epigenome, and most of the adaptation to environmental changes involves these cellular components. The current human genome is the result of a long evolutionary process, where environment-induced mutations have led to highly polymorphic DNA sequences. Also, acting upon the genome, the environment can select previously established genotypes. For example, in a population heavily exposed to HDMs, the possibilities of finding genotypes predisposing to high-IgE responsiveness are greater and then the prevalence of HDM sensitization will increase. This is probably what has occurred in the tropics, where HDM exposure is high and permanent, but could happen in temperate zones if HDM exposure increases by the reasons describe above (Fig. 2).
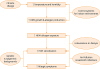 | Fig. 2A general and simplistic view of multiple factors that could affect HDM growth, allergen exposure, sensitization and allergic symptoms. The final outcomes depend on the interaction between genetic, environmentally protective and risk factors. Although some environmental changes are global, the effects on HDM sensitization and symptoms in the near future are expected to be regional.HDM, house dust mite.
|
Several studies support a genetic predisposition to asthma117 and also to get IgE sensitized to HDM allergens.118 The susceptibility loci to HDM sensitization include alleles in the major histocompatibility complex (MHC) and other out-of-MHC genes.119120121 Since the distribution of susceptibility alleles differ among human populations, we may expect that some groups will be more affected by global changes in HDM exposure. Migration studies have revealed that East Asian populations may be more susceptible to allergy and especially to HDM sensitization.6122 Some African-descent populations have also shown susceptibility alleles for increased IgE levels123 that are of relevance in HDM responses.124 It is thought that global environmental changes will modify the distribution and allergenicity of HDMs, which may increase HDM sensitization by allowing new contexts of HDM exposure. These increased rates will be in part explained by individuals with an “underlying” predisposition that become newly-HDM sensitized. On the other hand, there are protective alleles that may render some individuals non-affected by new HDM exposure. The context of exposure will also be critical in determining susceptibility because mite allergen levels mediate gene-environment interactions.125 For instance, the association between polymorphisms in the IL4 gene and HDM sensitization depended on Der p 1 allergen levels in carpet dust samples.126
Since the epigenome is more susceptible to environment-induced modifications, particularly those induced by air pollution, it is expected that recent environmental changes could exert part of their effects through epigenetic modification. Epigenetic mechanisms have been associated with the pathogenesis of allergic diseases and IgE sensitization. Experimental work on HDM-induced epigenetic modifications has revealed several alterations in the bronchial tissue leading to inflammation and bronchial hyperreactivity. These studies have shown that the epigenome might influence the susceptibility to mite sensitization by modifying DNA methylation in B cells _ENREF_122,127 and the hypomethylation of the IL13 gene.128 More interestingly, HDMs can induce epigenetic modifications in mouse models of airway inflammation, changing the methylation pattern of important genes such as PDE4D129 and TGFB1.130 Whether these effects are consequence of the allergen-induced immune response or a direct action of HDM allergens remain to be defined; however, using an ex vivo model of inflammation in human bronchial epithelial cells, HDMs induce the same epigenetic modifications as does diesel exhaust,131 suggesting that HDMs, in addition to inducing IgE-mediated bronchial inflammation, can alter the epigenetic patterns of cells involved in bronchial homeostasis, inducing inflammation. Hence, environmental exposure affecting the epigenome or polymorphisms influencing the interaction between the genome and the epigenetic machinery may play a role in modulating the gene-environment signals that lead to mite sensitization. In addition, epigenetic modifications might be inherited by transgenerational mechanisms,132 which means that they could be accumulated among populations.133 The process of human adaptation to the multiple changes of environment is difficult to predict and deserves further research on many fields. This fundamental interrogate has started to be analyzed in bird populations.134
CONCLUSION
This review shows that recent environmental changes are probably affecting the level and frequency of HDM exposure, which could partially explain the increase in HDM sensitization and asthma symptoms in some countries and regions. Further epidemiological and experimental research is needed to confirm these effects and to design prevention programs. However, adequate control of human-induced harmful environmental changes is essential for stopping the increasing trends of allergic diseases.
 XML Download
XML Download