

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Non-alcoholic fatty liver disease (NAFLD) associated with excessive lipid accumulation in the liver is the most common cause of chronic liver diseases, including obesity, dyslipidemia, hypertension, insulin resistance, and diabetes [1]. At the molecular level, NAFLD involves epigenetic alterations in the liver [2]. Lysine (Lys) acetylation is a crucial post-translational modification for the epigenetic regulation of genes. At least 2000 proteins in human cells can be post-translationally modified at lysine residues via acetylation [34]. Nearly 65 percent of these Lys-acetylated proteins are correlated with metabolism [567]. Thus, the dysfunction of Lys acetylation is major cause of metabolic syndrome, including obesity, cardiovascular disease, hypertension, type 2 diabetes, dementia, cancer, and NAFLD [89].
Two important families of enzymes, histone acetyltransferases (HATs) and histone deacetylases (HDACs), coordinate the steady state of acetylation [10]. HATs add acetyl groups to the Lys ε-amino residues of histones besides other non-histone proteins; in contrast, HDACs remove acetyl groups [1112]. To respond to changes in metabolic demands and energy flux, metabolic enzymes (non-histone proteins) alters their catalytic activity or stability through the acetylation [1314]. With respect to the regulation of metabolic enzymes in the liver, the balance of histone acetylation/deacetylation by HATs and HDACs is important.
NAFLD is conventionally explained by a “two-hit” model. Hepatic lipid accumulation is the first hit, causing insulin resistance, and excess inflammation by inflammatory cytokine interactions acts as the second hit [2]. However, recently, a new model in which multiple parallel hits are responsible for the development of NAFLD has been described [15]. The p300 HAT enzyme, which functions as a transcriptional coactivator, is involved in the first hit and second hit. It is co-recruited with nuclear factor-κB to regulate various inflammatory signaling pathways [16] and increases the transcriptional activity of carbohydrate-responsive element-binding protein (ChREBP), which is a coactivator of lipogenic genes via the acetylation of Lys-672 in the development of NAFLD [17]. However, little is known about the relationship between HATs and the development of NAFLD.
Unlike genetic changes, epigenetic modifications, like Lys acetylation, are potentially reversible and can be modified by environmental, dietary, and lifestyle factors [18]. Additionally, these epigenetic factors are closely associated with the phenotype and determine progression of NAFLD [19]. In this study, we investigated the correlation between HAT activity and protein acetylation in NAFLD and identified natural extracts (NExs) that attenuate lipid accumulation by modulating HAT activity in an NAFLD model of HepG2 cells.
MATERIALS AND METHODS
Cell culture
cells Human hepatocellular carcinoma (HepG2) cells were purchased from the American Type Culture Collection (Manassas, VA, USA) and cultured in a humidified atmosphere of 5% CO2 at 37℃ using high-glucose Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS) and antibiotics (Welgene, Daegu, Republic of Korea). To establish an NAFLD model in HepG2 cells, a non-fat BSA-conjugated combination of oleic acid (OA) (Sigma-Aldrich, St. Louis, MO, USA) and palmitic acid (PA) (Sigma) was used at a ratio of 4:1 (OPA). HepG2 cells were simultaneously treated with 200 µg of natural extracts and OPA for 24 h.
Preparation of natural extracts
Based on 2015 Korea National Health and Nutrition Examination Survey, about 80 natural food which are frequently consumed by Korean chosen, and 22 natural food extracts among them in which experiments had been first finished were analyzed in the current study (Table 1). The natural food used in this study purchased from a local market (Seongnam, Gyeongggi-do, Korea). A voucher specimen has been deposited in Korea Food Research Institute (KFRI). Following cleaning of specimens those were extracted in a 10-fold volume of 70% ethanol by shaking for 24 h at 25℃. Finally, the precipitate was eliminated by centrifugation at 8,000 × g for 30 min, and supernatants were lyophilized in a freeze- drier (II Shin, Dongdochum-Si, Korea).
Cell toxicity
Cells (approximately 5 × 104) were seeded in 24-well plates. After reaching approximately 70% confluence, cells were expos in OPA in presence or absence of NExs. After 24 h of incubation, 10 µL of WST-1 solution (Enzo Life Sciences, Inc., Farmingdale, NY, USA) was added for 3 h, and then absorbance of 100 µL of the supernatant was measured at 450 nm (Molecular Devices, Sunnyvale, CA, USA).
Oil red O staining
Cells (approximately 5 × 104/well) were seeded. Reached approximately 70% confluence, cells were treated in OPA, alone or in combination with NExs at 200 µg/mL. To measure lipid accumulation in HepG2 cells, we performed Oil Red O staining as previously described [20].
HAT assay
HeLa cell nuclear extract was purchased from Biovision Biotechnology (Milpitas, CA, USA). Nuclear extracts were assessed for HAT activity using a commercially available kit, according to the manufacturer's protocol (Biovision Biotechnology).
Histone extraction
In brief, 5 × 106 cells were seeded in 10-mm dishes. OPA was added for 18 h when the confluence reached approximately 70% (~2 × 108 cells). HepG2 cells were lysed according to the manufacturer's protocol (Abcam, Cambridge, MA, USA) pre-lysis buffer to isolate the nuclear fraction. The nuclear pellet was resuspended in lysis buffer, incubated for 30 min on ice, and centrifuged at 10,000 rpm for 5 min at 4℃. The supernatant was transferred to a new tube and DTT buffer was added. Samples were stored at −70℃ until analysis.
Quantitative real-time RT-PCR
Cells were seeded at 5 × 104 cells/well. After reaching approximately 70% confluence, cells were treated with or without OPA for 18 h. To observe the effect of HATs on lipid accumulation in HepG2 cells, C-646, a p300/CBP-specific inhibitor, was added with OPA to HepG2 cells for 18 h, and then we performed qRT-PCR as previously described [20]. The primers used in this study were listed up in Table 2.
Immunoblotting
Following treatment, cell extracts were supplemented with lysis buffer (Cell signaling) containing protease and phosphatase inhibitors (Roche, Basel, Switzerland) for 30 min and centrifugation at 20,000 × g for 20 min at 4℃. The cell lysates were separated by SDS-PAGE and transferred to nitrocellulose membranes. The membranes were incubated with specific antibodies as indicated in Table 3 overnight at 4℃, followed by incubation with a secondary antibody (Thermo Scientific, Rockford, IL, USA). Proteins were visualized using a ChemiDoc system (Bio-Rad) with an enhanced chemiluminescence substrate (Thermo Scientific).
Statistical analysis
qRT-PCR data were analyzed using t-tests. Correlations between HAT activity and lipid accumulation in HepG2 cells were evaluated by Pearson's correlation coefficients. Statistical analyses were carried out in SPSS (Ver. 20; SPSS Inc., Chicago, IL, USA). P < 0.05 was considered statistically significant.
RESULTS
The acetylation status of non-histone and histone proteins increased by OPA
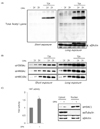
To examine whether the acetylation level of non-histone proteins and histone proteins increased in the NAFLD model, the acetylation status was evaluated in HepG2 cells with or without OPA treatment. In particular, cells were treated with 400 µM of OA and 100 µM of PA (1×) or 800 µM of OA and 200 µM of PA (2×) with or without trichostatin A (TSA). As shown in Fig. 1A, the acetylation level of total proteins was higher in the samples treated with OPA and TSA than in the control group. Additionally, the acetylation level was relatively higher in the group treated with a high concentration of OPA. Next, to observe changes in the specific acetylation status of histone tails, H3K9, H4K8, and H4K16, related to the expression of genes related to lipid accumulation in liver, were extracted from HepG2 cells. Unlike total protein acetylation, H3K9, H4K8, and H4K16 were acetylated without TSA treatment and the acetylation levels increased in proportion to the concentration of OPA (Fig. 1B). Although the levels of acetylation at H3K9, H4K8, and H4K16 increased in the groups treated with both OPA and TSA, similar to the patterns observed for the groups treated with OPA only, the degree of acetylation was much greater than that of groups treated with OPA only (Fig. 1B). Based on the close association between protein acetylation and HAT activity, we investigated whether HAT activity was greater in the NAFLD model than in the control. As expected, HAT activity was much greater in the group treated with OPA (Fig. 1C). Taken altogether, these results showed that the acetylation levels of both non-histone and histone proteins were positively related to HAT activity, which increased by lipid accumulation in HepG2 cells.
Lipogenic gene expression decreased by blocking HAT activity using C-646, a p300/CBP-specific inhibitor
We next investigate the effect of OPA on the expression of lipogenesis-related genes, such as peroxisome proliferator-activated receptor-γ (PPARγ), sterol regulatory element-binding protein-1c (SREBP1c), ATP citrate lyase(ACLY), and fatty acid synthase (FASN), in HepG2 cells. Consistent with the results of previous studies, we found that an increase in lipogenic gene expression was related to lipid accumulation (Fig. 2A). To elucidate the relationship between the mRNA expression of lipogenic genes and HAT activity, we incubated HepG2 cells with or without C-646, a specific p300/CBP inhibitor, in combination with OPA for 18 h. As shown in Fig. 2B, the mRNA expression of lipogenic genes, i.e., PPARγ, SREBP1c, ACLY, and FASN, was abrogated by blocking p300/CBP activity using C-646. Collectively, our data suggest that there is a correlation between the expression of lipogenic genes and HAT activity.
EMA and ESC effectively prevent OPA-induced lipid accumulation in HepG2 cells
We hypothesized that natural compounds that can block OPA-induced lipid accumulation may show relatively high anti-HAT activity. Thus, we selected 22 commonly consumed vegetables in Korea and measured their ability to prevent OPA-induced lipid accumulation in HepG2 cells. 200 µg/mL extract with OPA were treated in cells, and lipid accumulation was measured by Oil Red O staining (Fig. 3A, upper panel). To quantify it, absorbance of the dissolved Oli Red O dye in isopropanol was measured (Fig. 3A, lower panel). Lipid accumulation was greater in OPA-treated cells than in the control group, and most extracts, including the extract of Allium tuberosum (EAF), had a protective effect on lipid accumulation. In particular, the extract of Morus alba (EMA) and that of Schizandra chinensis Bail (ESC) reduced lipid accumulation by almost 30% compared to that in cells treated with OPA alone. The extracts of Platycodon grandiflorus, Fragaria x ananassa, Allium monanthum, and Malus domestica did not have preventive effects on OPA-induced lipid accumulation in HepG2 cells. We examine whether the effect of NExs on the inhibition of lipid accumulation occurred as cytotoxicity. HepG2 cells were incubated in the presence or absence of OPA, with or without NExs. As shown in Fig. 3B, NExs did not influence the viability of cells. Altogether, NExs used in this study significantly abrogates OPA-induced lipid accumulation without cytotoxicity.
ESC shows the strongest effect on HAT activity
To analyze the correlation between lipid accumulation and HAT activity, we used a cell-free system. NEs at the same concentrations used in the previous experiment were incubated with HeLa nuclear extract, and epigallocatechin-3-gallate (EGCG), which is a well-established specific HATi [21], was used for comparison. As shown in Fig. 4, anti-HAT activity was observed in 22 NEs. Among these, 10 NEs, i.e., EAT, EMA, ESC, ESO, ESL, ECU, EPF, EHT, EZO, and EPT, inhibited HAT activity by over 20% in comparison with activity in the control group (gray bars in Fig. 4). ESC demonstrated the strongest anti-HAT effect, and EMA also showed a relatively strong inhibitory effect, supporting the correlation between lipid accumulation and HAT activity.
Lipid accumulation and HAT activity are positively correlated
To further evaluate the correlation between lipid accumulation and HAT activity, Pearson's correlation coefficients were calculated using Oil Red O absorbance data and HAT activity for each NE (Fig. 5). The correlation coefficient was 0.604 (P = 0.0035), suggesting that there is a positive correlation between lipid accumulation and HAT activity.
DISCUSSION
NAFLD is the most common liver disease worldwide. Many studies have focused on the effects of natural dietary compounds on lipid metabolism to develop new therapeutic strategies [22]. However, lifestyle influences the development and progression of NAFLD via epigenetic regulation [23], since it is a complex disease determined, in part, by environment factors [24]. Thus, epigenetic mechanisms involved in lipid metabolism are regarded as potential preventive or therapeutic targets [25].
Our data supported the results of previous studies showing that NAFLD may be controlled by epigenetic regulatory mechanisms. In particular, the balance between histone acetylation/deacetylation at the N-termini, which are catalyzed by HAT and HDAC, respectively [26], influences gene expression profiles in NAFLD [27]. As shown in Fig. 1B, histone H3K9, H4K8, and H4K16 acetylation increased dramatically following OPA treatment, and the acetylation status increased when HDAC activity was inhibited by TSA, an HDAC-specific inhibitor, indicating that aberrant histone acetylation by an imbalance between HAT and HDAC activity is involved in the hyperacetylation of histones, resulting in the development and progression of NAFLD. In adipocytes, histone H3K56 acetylation was first identified in a genome-wide profile analysis [28]. Mikula et al. demonstrated that the acetylation of histones H3K9 and H3K18 in the TNFα and Ccl2 promoters was increased in the livers of mice fed a high-fat diet [29]. Furthermore, our results indicated that the total protein acetylation status increased significantly in cells treated with OPA in combination with TSA. Unexpectedly, acetylated-Lys was not detected without HDAC inhibition in extracts of the total cell lysate, unlike that of histones (Fig. 1A), suggesting that HDACs, which are predominantly localized in the cytosol or shuttle between the cytoplasm and the nucleus, are closely involved in the regulation of acetylation on non-histone proteins related to lipid accumulation. Similarly, a previous study reported that total protein acetylation is significantly increased in the hearts of HFD-fed mice [30], suggesting that lipid accumulation-induced hyperacetylation is not limited to the liver and is involved in obesity-related diseases in various organs. In particular, SREBP1c is acetylated by p300/CBP in both cells and mouse livers, leading to increased stability and recruitment to its lipogenic target gene promoter [31]. p300/CBP hyperactivity was also associated with ChREBP acetylation and hepatic steatosis in a mouse model of obesity [17]. Accordingly, the increase in HAT activity in OPA-treated cells (Fig. 1C) was expected.
Transcription is a highly regulated process, and acetylation plays a major role in this regulation [32] by the various conformational changes of the chromatin, including charge neutralization [33], consequently providing unique biding surfaces for activators of transcription [34]. PPARγ and SREBP1c regulate genes required for lipid metabolism, such as ACLY or FASN [35]. The mRNA expression of PPARγ is up-regulated in the liver of obese patients with NAFLD, reinforcing the lipogenic mechanism in response to SREBP-1c induction in the development of NAFLD [36]. Considering previous studies, our data (Fig. 2A) showing the increase in lipogenic gene expression is not unexpected. However, the reduction in lipogenesis-related gene transcription following C-646 treatment was noteworthy, indicating that dynamic histone acetylation and deacetylation are also involved in the transcriptional activation of lipogenesis-related genes.
Currently, there are no approved pharmacological agents or FDA guidelines for the treatment of NAFLD [37]. Anti-obesity drugs, such as orlistat and sibutramine, are used for NAFLD but have potentially hazardous side effects [38]. Accordingly, a variety of natural phytochemical compounds to prevent NAFLD have been explored. In our study, 15 of 22 NExs prevented OPA-induced lipid accumulation in HepG2 cells (Fig. 3A). The anti-lipogenic effects of the extracts, including EMA, ESC, and EFA, are well established [3940]. However, our aim was to elucidate the mechanisms involved in the beneficial effects of NExs on NAFLD prevention from a new perspective, i.e., epigenetics. Alterations in acetylation status associated with chronic metabolic diseases, including the abnormal regulation of hepatic lipid metabolism, have been related in the development and progression of NAFLD [2]. Certain bioactive foods may have a beneficial effect on NAFLD via their effects on acetylation. Interestingly, two NExs, EMA and ESC, demonstrated strong anti-lipogenic effects and inhibited HAT activity (Fig. 4), suggested that there is a positive correlation between lipid accumulation and HAT activity. An analysis of the correlation between these two factors supported this finding. Although the relationship was not strong (Fig. 5, Pearson's correlation coefficient = 0.604), it was sufficient to suggest a new approach for the prevention or treatment of NAFLD based on natural foods with epigenetic effects.
The present study had many limitations. The number of preventative natural food extracts examined in the study was small. To definitively establish the correlation between lipid accumulation and HAT activity, further studies of the molecular mechanism are needed.







 XML Download
XML Download