

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Kawasaki disease is a disease in children with acute vasculitis whereby the blood vessels throughout the body are inflamed.12 The incidence of Kawasaki disease varies by race and country, but the highest incidence is observed in Asian countries, especially in Japan and Korea.34 The cause and pathological mechanism of Kawasaki disease remain largely unknown. Moreover, a molecular marker for this disease has not been established. Current criteria to diagnose Kawasaki disease are mainly clinical symptoms including polymorphous rash, conjunctival injection, strawberry tongue, cervical lymphadenopathy, and desquamation.5 Because of the high probability of coronary artery aneurysms and other serious conditions without prompt treatment, establishing a molecular marker to quickly diagnose Kawasaki disease is essential.1 Therapy for Kawasaki disease includes anti-inflammatory treatment using intravenous immunoglobulin (IVIG) with aspirin and/or steroids such as prednisone, although the detailed mechanisms for these treatments are not well understood.25 Thus, more studies are needed to determine the pathology and identify specific biomarkers for Kawasaki disease.
Circular RNA (circRNA) is a class of single-stranded RNA with a closed circular structure. In humans, over ten thousand circRNAs have been discovered, and most do not yet have annotated functions.6 These circRNAs originate from diverse sources, but most are produced from the exons of host genes through back-splicing.78 During the back-splicing reaction, the 3ʹ end of a downstream exon is connected into the 5ʹ end of an upstream exon, producing the closed circular structure. The working mechanism of several circRNAs has been reported to date, where it has been shown that many circular RNAs have a microRNA (miRNA)-binding site in their sequences.9 Because miRNAs primarily suppress the translation of their target mRNAs, circRNAs can function to indirectly increase the translation of protein-coding mRNAs through absorbing miRNAs. Other studies have shown that circRNAs repress the function of proteins by binding them.10 In addition, some circRNAs have an open reading frame that is able to produce small proteins.1112 Therefore, circRNAs can behave as important regulators in cells.
Although the function of circRNAs has been reported in human pathogenesis, the role of circRNAs in Kawasaki disease has not yet been reported. To understand how differential circRNA expression may affect the regulatory mechanism of this disease, circRNA expression profiles were analyzed using publicly available datasets from studies that performed RNA sequencing on coronary artery tissues from patients with Kawasaki disease. Several circRNAs were identified to be differentially expressed in untreated Kawasaki disease tissues, but their level returned to normal after treatment. The data obtained from this study will be useful for future studies focusing on the underlying molecular mechanism of Kawasaki disease progression and for determining molecular markers for this disease.
MATERIALS AND METHODS
1. Samples used for analysis
RNA sequencing data from coronary artery tissue samples from controls and patients with Kawasaki disease were obtained from the Gene Expression Omnibus (GEO) dataset GSE64486. The information for patients with Kawasaki disease and controls was described previously.13 Briefly, the control group had normal coronary artery histology. Among the two groups of patients with Kawasaki disease, the untreated group did not receive IVIG therapy after the onset of fever while the treated group received treatment with IVIG, aspirin, steroids, and/or infliximab.13
2. Analysis of circRNA expression from RNA sequencing data
The raw data produced from RNA sequencing was filtered using the Trimmomatic trimmer tool to trim the 5ʹ and 3ʹ ends of sequences and to remove reads with overall low quality (option: LEADING: 5 TRAILING: 5 SLIDINGWINDOW: 4:20 MINLEN: 36).14 The remaining adaptor sequences were also trimmed during this procedure. The filtered sequences were aligned to the human genome using the STAR aligner with paired reads or single reads, respectively.15 Using the alignment information, circRNA expression was calculated using the DCC algorithm, which resulted in the identification of 9,134 circRNAs.16 Among them, only 311 circRNAs with average counts greater than 2 among whole samples were selected for further analysis. For the selected circRNAs, their expression levels were normalized by dividing the counts with the entire circRNA counts in each sample, and then the fold change of circRNA expression was calculated among the groups. The host gene and exon structures where the circRNAs originated were inspected manually using the UCSC Genome Browser server, and only those circRNAs whose exon structures fully overlapped with the exon structure of the host genes were selected.17
3. Analysis of circRNA regulation on miRNAs
To identify potential miRNAs that may be regulated by the selected circRNAs, the miRNA list reported to be differentially expressed between artery tissues from patients with Kawasaki disease and those from control patients was obtained.18 There were 26 miRNAs on this list, including miR-92a, miR-93, miR-127-3p, miR-146b-5p, miR-150, miR-181b, miR-185, miR-210, miR-223, miR-320a, miR-320b, miR-423-5p, miR-548c-5p, miR-566, miR-638, miR-650, miR-654-5p, miR-663, miR-663b, miR-675, miR-1246, miR-1274b, miR-1285, miR-1291, miR-1300, and miR-1307. The binding sites for these miRNAs in the circRNA sequences were analyzed using the TargetScan algorithm.19
RESULTS
To identify circRNAs that may be involved in the pathogenesis of Kawasaki disease, RNA sequencing data from coronary artery tissue from two groups of patients with Kawasaki disease and a group of control patients was utilized (Fig. 1A).13 The control group contained seven samples with normal coronary artery histology. The two patient groups were classified into untreated Kawasaki patients and Kawasaki patients treated with IVIG, with each group containing four samples. After trimming low-quality reads from the sequencing results, circRNA expression levels were calculated using the DCC algorithm, which counts the chimeric reads produced by linking the 3ʹ end of a downstream exon and the 5ʹ end of an upstream exon. Because one control sample and one sample from the untreated Kawasaki disease group had a small number of circRNA counts, these samples were removed from the subsequent analysis (Fig. 1A).
Fig. 1
Identification of circRNAs expressed in coronary artery tissue from control and Kawasaki disease groups. (A) The procedure used to identify circRNAs in the control and Kawasaki disease samples. Based on data from the GSE64486 dataset, the expression level of circRNAs in each group was calculated using the DCC algorithm and differentially expressed circRNAs were selected for further study.1316 (B) Average expression counts of highly expressed circRNAs. circRNAs with normalized average counts greater than 10 are shown. (C) The number of circRNAs produced from each gene was calculated.
circRNA, circular RNA.

Overall, 311 circRNAs with reliable expression counts were identified in the coronary artery tissues (Supplementary Table 1, see Methods). Among these circRNAs, circCDR1 exhibited the highest expression in the samples (Fig. 1B). This circRNA was previously reported to be a potent miRNA suppressor with over 60 binding sites for miR-7.2021 In most cases, only a single circRNA type was produced from each host gene, but in some cases, diverse circRNAs were produced from a different combination of host gene exons (Fig. 1C).
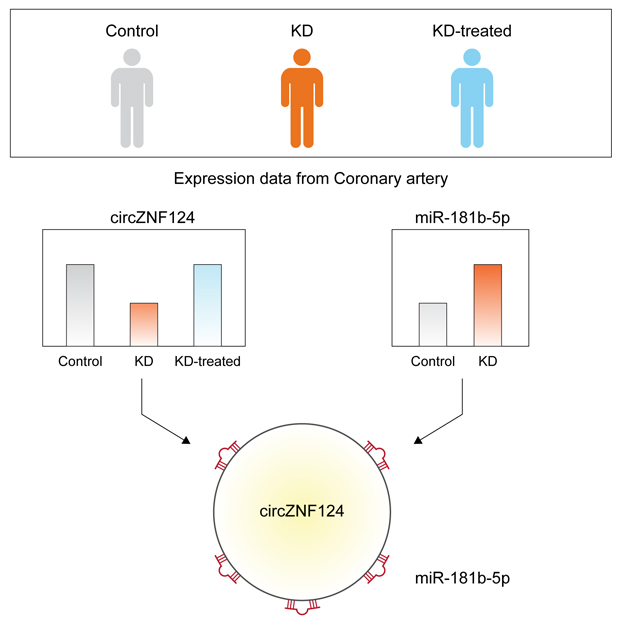
To identify circRNAs that may have a role in the progression of Kawasaki disease or may be used as a potential biomarker, the fold change in expression of each circRNA was calculated and circRNAs with differential expression between any two groups were selected. This analysis resulted in 32 circRNAs (Supplementary Table 2). Among the circRNAs that were differentially expressed between the control group and the untreated Kawasaki disease group, five circRNAs including circHIPK3, circZNF124, circWHAMMP1, circSLAIN2, and circATM exhibited recovered expression after treatment (Fig. 2).
Fig. 2
Selected circRNAs with probable roles in Kawasaki disease. Among the circRNAs listed in Supplementary Table 2, only circRNAs that showed differential expression between control and untreated Kawasaki disease groups (p<0.05), and between untreated and treated Kawasaki disease groups (p<0.05), but no significant difference between the control and treated Kawasaki disease groups (p>0.05), were selected. p values were calculated by a t-test between samples from 2 groups, respectively.
circRNA, circular RNA.
*p<0.05; †p<0.01.

CircRNAs are generally involved in gene regulatory networks by suppressing the function of miRNAs. In a previous study, miRNA expression levels were compared between coronary artery samples from patients with Kawasaki disease and control patients, and 26 miRNAs were reported to show significantly increased expression in the Kawasaki disease group.18 Because expression of the selected circRNAs was decreased in the untreated Kawasaki disease group compared to that in the control group (Fig. 2), it was predicted that the target miRNAs of these circRNAs would be increased in the same group. Using TargetScan, a miRNA target prediction algorithm, the target sites of the increased miRNAs in the selected circRNA sequences were predicted (Fig. 3A).19 This analysis identified that circWHAMMP1 and circZNF124 are most likely regulated by miR-663b and miR-181b-5p, respectively (Fig. 3B and C). Strikingly, there were five binding sites for miR-181b-5p in the sequence of circZNF124, suggesting that circZNF124 may be an efficient suppressor of miR-181b-5p.
Fig. 3
Possible working mechanism of circRNAs as miRNA suppressors. (A) Analysis of circRNAs as miRNA suppressors. Among the increased miRNAs in patients with Kawasaki disease, miRNAs with predicted binding sites in circRNA sequences were selected using TargetScan.19 (B-C) The miRNA binding sites in circRNAs are depicted for (B) circWHAMMP1 and (C) circZNF124. (D) Base pairing between the predicted miRNA binding sites and circRNAs is shown. The nucleotide position of the back-splice junction depicted in (B, C) was set as zero, and the relative position of the nucleotide sequences was calculated.
circRNA, circular RNA; miRNA, microRNA.
DISCUSSION
In this study, the expression profile of circRNAs in the artery tissue of patients with Kawasaki disease was analyzed using RNA sequencing data from the GEO database.1322 By comparing the expression level of circRNAs among control patients, patients with untreated Kawasaki disease, and treated patients with Kawasaki disease, five circRNAs were identified that could be used as molecular biomarkers to reflect the status of Kawasaki disease (Fig. 2). Among those circRNAs, only the function of circHIPK3 has been studied previously. This circRNA was previously reported to be involved in vascular dysfunction of the retina in patients with diabetes.23 Moreover, another study showed that circHIPK3 is involved in regulating human cell growth by suppressing multiple miRNAs, suggesting that circHIPK3 is an important regulator in human diseases.24
To date, many studies have identified the function of miR-663b and miR-181b-5p, which may be suppressed by circWHAMMP1 and circZNF124, respectively. One previous study found that miR-663b increases cell proliferation and migration in nasopharyngeal carcinoma.25 Another study showed that miR-181b expression decreases during the aging process and is involved in regulating vascular stiffness.26 Although there is still no direct evidence that these miRNAs have any role in the progression of Kawasaki disease, misregulation in miRNA activity due to altered circRNA expression may affect cellular pathways in arteries. Thus, it will be interesting to identify the role of these miRNAs in the progression of Kawasaki disease in future studies.
 XML Download
XML Download